Floem

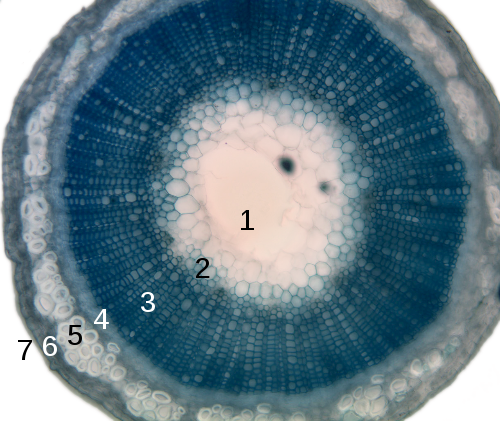
- Srž
- Protoksilem
- Metaksilem
- Floem
- Sklerenhim (vlakno like)
- Kora
- Epiderma
Floem (grč. φλοıός - floiόs = kora) ili lika (liko) zajednički je naziv za mreže provodnih snopova u provodnim sudovima viših biljaka. Služi za transport organskih tvari koje su nastale u procesu fotosinteze u listovima i ostalim organima sa hlorofilom, za razliku od ksilema koji služi za provođenje vode s rastvorenim mineralnim tvarima[1] do dijelova gdje su potrebni. Taj se proces transporta naziva translokacija.[2]
Ovaj termin uveo je Nägeli, 1858.[3][4]
Struktura
[uredi | uredi izvor]
Floemsko tkivo se sastoji od ćelija koje provode, uglavnom se nazivaju sitaste parenhimske ćelije, uključujući i specijalizirane ćelije za pridruživanje ili albuminozne ćelije i nespecijalizirane ćelije i potporne ćelije, kao što su vlakna i sklereidi.[5]
Provodne ćelije – sitasti elementi
[uredi | uredi izvor]
Sitasti elementi su vrsta ćelija koje su odgovorne za transport šećera kroz biljku.[5] U zrelosti im nedostaje jedro i imaju vrlo malo organela, pa se za većinu svojih metaboličkih potreba oslanjaju na ćelije pratilice ili albuminozne ćelije. Ćelije sitastih cijevi sadrže vakuole i druge organele, poput ribosoma, prije nego što sazriju, ali one uglavnom prelaze na ćelijski zid i rastvaraju se u zrelosti; to osigurava da mogu malo ometati kretanje tečnosti. Jedna od rijetkih organela koju sadrže u zrelosti je grubi endoplazmatski retikulum, koji se može naći u plazma membrani, često u blizini plazmodezmi što ih povezuje sa svojim pratilicama ili albuminoznim ćelijama. Sve sitaste ćelijie imaju krajeve pora na svojim krajevima koji rastu iz modificiranih i proširenih plazmodezmi, nazvanih sitasta područja. Pore su pojačane pločicama polisaharida zvanim kaloza.[5]
Funkcija
[uredi | uredi izvor]Za razliku od ksilema (koji se sastoji uglavnom od mrtvih ćelija), floem čine još žive ćelije koje prenose biljne sokove. Sok je otopina na vodenoj osnovi, ali bogata šećerom stvorenim u fotosintezi. Ti se šećeri transportiraju se u nefotosintetske dijelove biljke, poput korijena, ili u skladišne strukture, poput gomolj ili lukovica. Tokom perioda rasta biljke, obično tokom proljeća, organi skladištenja poput korijena su izvor šećera, a mnoga područja biljke ima ga u rezervi. Kretanje u floemu je višesmjerno, dok je u ćelijama ksilema jednosmjerno (prema gore).
Nakon perioda rasta, kada su meristemi u stanju mirovanja, listovi su izvori, a organi za sodlaganje su skladišta. Razvijaju sjemenosne organe (poput ploda) su uvijek skladišta. Zbog ovog višesmjernog protoka, zajedno s činjenicom da se sok ne može lako kretati između susjednih sitastih cijevi, nije neobično da sok u susjednim sitastim cijevima struji u suprotnim smjerovima.[6] Dok se kretanje vode i minerala kroz ksilem uglavnom omogućava negativni pritisak (napetost), kretanje kroz floem pokreće se pozitivnim hidrostatskim pritiscims. Ovaj se postupak naziva translokacija (premještanje), a provodi se procesom koji se zove floemski utovar i istovar.
Smatra se da i floemski sok ima ulogu u slanju informativnih signala kroz vaskularne biljke. Obrazaci utovara i istovara u velikoj mjeri određeni su vodljivosti i brojem plazmodezmi i funkcijom rastvora, što ovisi o položaju rastvora – specifičnost plazma membrane i transportnih proteina. Nedavni dokazi govore da su pokretni proteini i RNK dio biljnog komunikacijskog signalizacijskog sistema na daljinu. Postoje i dokazi za usmjereni transport i sortiranje makromolekula tokom prolaska, preko plazmodezmi.[7] Organske molekule, poput šećera, aminokiselina], određenih hormona], pa čak i iRNK prenose se u floem kroz elemente sitastih cijevi.[7]
Opojasivanje
[uredi | uredi izvor]Budući da se u većini biljaka floemske cijevi nalazi izvan ksilema, drvo ili druga biljka mogu se ubiti prstenastim uklanjanjem kore na deblu ili stabljici. Sa uništenim floemom, hranjive tvari ne mogu doći do korijena, a drvo/biljka će uginuti. Drveće koje se nalaze u područjima sa životinjama poput dabra su ranjiva, jer dabrovi žvaču koru na prilično preciznoj visini. Ovaj postupak je poznat kao pojasni i može se koristiti u poljoprivredne svrhe. Naprimjer, ogromne količine voća i povrća viđenog na sajmovima i karnevalima proizvedeno je preko pojasa skinute kore. Poljoprivrednik bi ogulio pojas u podnožje velike grane i uklonio sve voće / povrće osim na jednoj grani. Dakle, svi šećeri proizvedeni od lišća na toj grani nemaju spremište ugljen-dioksida. Voće / povrće koje se na taj način raste u mnogostruko većoj mjeri od svoje normalne veličine.
Porijeklo
[uredi | uredi izvor]Kada je biljka embrionskoj fazi, vaskularno tkivo nastaje iz tkiva prokambija, koji se nalazi u središtu embriona. Protofloem se pojavljuje u sredini provodnog suda koji se proteže u kotiledonski čvor, što predstavlja prvo pojavljivanje lista u angiospermi, koje formiraju kontinuirane provodne cijevi. Hormon auksin koji prenosi protein PIN1 odgovoran je za rast tih protofloemskih niti, što signalizira konačan identitet tih tkiva. U tom procesu također učestvuju iSHR i mikroRNK165 / mir-166 mikroRNK prekursor, dok kaloza sintaza 3 (CALS3) inhibira lokacije na koje mogu ići SHR i mikroRNK165. U embrionu, korijenski floem se neovisno razvija u gornjem hipokotilu, koji leži između korijena embriona i kotiledona.[8] Kod odraslih biljaka, lika potiče i raste prema van iz meristemskih ćelij vaskularnog kambija. Floem se proizvodi u fazama. Primarni floem je postavljen apikalnim meristemom i razvija se iz prokambija. Sekundarni nastaje od vaskularnog kambija u unutrašnjost uspostavljenog sloja (slojeva) floema. U nekim porodicama eudikotiledona (Apocynaceae, Convolvulaceae, Cucurbitaceae, Solanaceae, Myrtaceae, Asteraceae, Thymelaeaceae), floem se također razvija na unutrašnjoj strani vaskularnog kambija; u ovom slučaju pravi se razlika između "vanjskog (eksternog) floema i "unutrašnjeg (internog) floema" ili "intraksilarnog floema. Unutrašnji floem uglavnom je primarni i započinje diferencijaciju kasnije od vanjskeog i protoksilema, iako to nije bez izuzetaka. U nekim drugim porodicama (Amaranthaceae, Nyctaginaceae, Salvadoraceae), kambij također periodično formira unutrašnje niti ili slojeve floema, ugrađene u ksilem: takve niti floema nazivaju se uključeni floem ili interksilarni floem.[9]
Upotreba
[uredi | uredi izvor]
Floem borovih stabala korišten je u Finskoj i Skandinaviji kao zamjena za hranu u doba gladi, pa čak i u dobrim godinama na sjeveroistoku. Snabdevanje floemom iz prethodnih godina pomoglo je da se ublaži velika glad u periodu 1866-1868. Floem se sušio i mljeo u brašno i miješao sa raži, kako bi se tvrdi napravio tamni hljeb (kruh). Najmanje se cijenio „svileni“, hljeb napravljen bez pravog raženog ili žitnog brašna. Nedavno je „pettu“ ponovo postao dostupan zbog znatiželje, a neki su tvrdili da daje i zdravstvene beneficije. Međutim, njegov sadržaj energije u hrani je nizak u odnosu na raž ili druge žitarice.
U Bosni i Hercegovini i okolnim zemljama, likovina još uvijek ima mnogostruku opotrebu, kao organsko vlakno za vezanje, uvezivanje i pletenje u različitim prilikama, naprimjer u kalemarstvu, korparstvu i raznim obrtima.
Vlakna lika dobijaju se vlaženjem ili ključanjem, te ljuštenjem kore drveća, kao što su lipa, brijest, vrba, bagrem i druga. Zbog svoje snage koriste se za vezanje u seoskim domaćinstvima i poljoprivredi, za pletenje i uvijanje užadi, izradu pletenih predmeta (prostirka, vreće, košare, roštilj, obuća, stolnjaci, zastori). Trake likovine odlišća nekih palmi poznate su kao rafija.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Lalonde S. Wipf D., Frommer W.B. (2004). "Transport mechanisms for organic forms of carbon and nitrogen between source and sink". Annu Rev Plant Biol. 55: 341–72. doi:10.1146/annurev.arplant.55.031903.141758. PMID 15377224.
- ^ Collins Edexcel International GCSE Biology, Student Book (ISBN 978-0-00-745000-8) p.124
- ^ Nägeli, Carl (1858). "Das Wachstum des Stammes und der Wurzel bei den Gefäßpflanzen und die Anordnung der Gefäßstränge im Stengel" [The growth of the stem and of the root among vascular plants and the arrangement of the vascular strands in the stalk]. Beiträge zur Wissenschaftlichen Botanik (Contributions to Scientific Botany) (jezik: German). 1: 1–156.CS1 održavanje: nepoznati jezik (link) From p. 9: "Ich will die beiden Partien Dauergewebe, welche von dem Cambium nach aussen und nach innen gebildet werden, Phloëm und Xylem nennen." (I will call the two parts of the permanent tissue, which are formed by the cambium outwardly and inwardly, "phloëm" and "xylem".)
- ^ Buvat, Roger (1989). "Phloem". Ontogeny, Cell Differentiation, and Structure of Vascular Plants. str. 287–368. doi:10.1007/978-3-642-73635-3_10. ISBN 978-3-642-73637-7.
- ^ a b c Raven, Peter H.; Evert, R.F.; Eichhorn, S.E. (1992). Biology of Plants. New York, NY, U.S.A.: Worth Publishers. ISBN 978-1-4292-3995-0.CS1 održavanje: upotreba parametra authors (link)
- ^ Canny, MJ. Phloem Translocation. str. 124.
- ^ a b Turgeon, Robert; Wolf, Shmuel (2009). "Phloem Transport: Cellular Pathways and Molecular Trafficking". Annual Review of Plant Biology. 60: 207–21. doi:10.1146/annurev.arplant.043008.092045. PMID 19025382.
- ^ Lucas, William, et al. The Plant Vascular System: Evolution, Development and Functions. Journal of Integrative Plant Biology. 55, 294-388 (2013) PubMed
- ^ Evert, Ray F. Esau's Plant Anatomy. John Wiley & Sons, Inc, 2006, pp. 357–358, ISBN 0-470-04737-2.
Vanjski linkovi
[uredi | uredi izvor]| Discipline | |
|---|---|
| |
| Rast i habitus (izgled) | |
| Razmnožavanje | |
| Taksonomija biljaka | |
| Primjena | |
| |