Int. J. Plant Biol. 2025, 16(1), 11; https://doi.org/10.3390/ijpb16010011 - 10 Jan 2025
Abstract
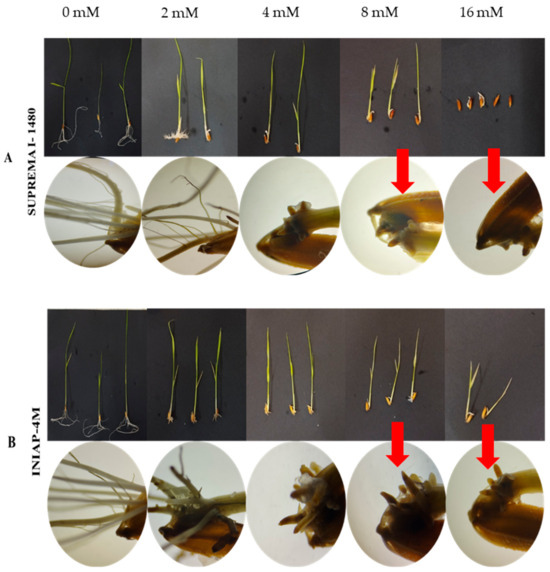
Moringa (Moringa oleifera Lam.) is a tree that grows in tropical and subtropical regions. In this study, the plants were grown in a temperate climate zone from seeds collected at the Island of St. Lucia. Cultivation was carried out in the field
[...] Read more.
Moringa (Moringa oleifera Lam.) is a tree that grows in tropical and subtropical regions. In this study, the plants were grown in a temperate climate zone from seeds collected at the Island of St. Lucia. Cultivation was carried out in the field and in a greenhouse in Prešov, East Slovakia. Leaf samples were taken from young plants and dried naturally. In the ethanol and hot water extracts of the leaves, the dry matter, total phenolic substances, and antioxidant activity were determined using three methods: superoxide anion radical scavenging activity, hydroxyl radical scavenging activity, and ferric reducing ability of plasma (FRAP) assay. The highest amount of total phenols was detected in the ethanolic extract of the leaves from the field 911.14 mg GAE L−1, resp. 69.70 mg GAE g−1 DM. The lowest amount was noticed in the leaves from the greenhouse 408.88 mg GAE L−1, resp. 13.07 mg GAE g−1 DM. The amount was significantly lower in the aqueous extracts. A high antioxidant activity of the leaves from the field was detected in all ethanolic and hot water extracts. Both types of leaf extracts from the greenhouse showed statistically significant lower antioxidant activity. The obtained results indicate that outdoor cultivation in a temperate climate zone was stressful for the plants, leading to an increased formation of phenolic substances, and consequently to higher antioxidant activity.
Full article
(This article belongs to the Section Plant Physiology)