Ubikvitin

| |||||||||
| Dijagram ubikvitina. Sedam lizinskih bočnih lanaca je obojeno narandžasto. | |||||||||
| Identifikatori | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Simbol | ubiquitin | ||||||||
| Pfam | PF00240 | ||||||||
| InterPro | IPR000626 | ||||||||
| PROSITE | PDOC00271 | ||||||||
| SCOP | 1aar | ||||||||
| Superfamilija | 1aar | ||||||||
| |||||||||
Ubikvitin je mali regulatorni protein, koji je prisutan u skoro svim tkivima eukariota. Ubikvitinacija je proces posttranslacione modifikacije nekog proteina, koja nastaje njegovim vezivanjem za jedan ili više ubikvitinskih monomera kovalentnim vezama.[1] Najčešća svrha ubikvitinacije je označavanje proteina za razgradnju. Ubikvitinska etiketa usmerava proteine ka proteasomu, koji je veliki proteinski kompleks u ćelijama koji razlaže i reciklira nepotrebne proteine. Osim toga, ubikvitinacija kontroliše stabilnost, funkciju i unutrašnji ćelijski položaj velikog broja proteina. Proces ubikvitinacije započinje enzim E1.[2]

Ubikvitin (prvotno sveprisutni imunopotentni polipeptid) je bio identifikovan 1975. godine kao sveprisutni protein mase 8.5 kDa. Osnovne uloge ubikvitina otkrivene su u ranim 1980-im u istraživanjima Foks Čejs instituta za rak, a otkrila ga je ekipa naučnika koju su sačinjavali Aron Ciechanover, Avram Hershko i Irvin Rose, koji su za svoje otkriće dobili Nobelovu nagradu za hemiju 2004. godine.[3][4][5]
Ubikvitinski sistem je prvotno okarakterisan kao ATP-zavisni proteolitički sistem prisutan u ćelijskim ekstraktima. Za termostabilani polipeptid prisutan u ovim ekstraktima, ATP-zavisni proteolitički faktor 1 (APF-1), je utvrđeo da je povezan kovalentnom vezom za model supstrata lizozima, u ATP i Mg2+ zavisnim procesima. Višestruki APF-1 molekuli su bili povezani s jednim molekulom supstrata izopeptidnim vezama. Ustanovljeno je da se njihovi konjugovani proteini brzo razlažu uz oslobađanje slobodnih APF-1. Ubrzo nakon toga, razjašnjena je konjugacija APF-1 proteina, koji je nazvan ubikvitinom. Karboksilna grupa aminokiseline glicina (Gly76) je otkrivena kao mjesto spajanja sa lizinom supstrata.
| Broj aminokiselina | 76 |
| Molekulska masa | 8564.47 Da |
| Izoelektrično polje (pI) | 6.79 |
| Nazivi gena | RPS27A (UBA80, UBCEP1), UBA52 (UBCEP2), UBB, UBC |
Ubikvitin je protein koji se sastoji od 76 aminokiselina i ima molekulsku masu od 8.5 kDa. Ključne karakteristike su C-terminalni rep i 7 lizina. Visoko je konzerviran među eukariotskim vrstama: ljudi i kvasci dele 96% sekvence. Sekvenca ljudskog ubikvitina, pri čemu svako slovo označava aminokiselinu, je:
| MQIFVKTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIFAGKQLEDGRTLSDYNIQKESTLHLVLRLRGG |
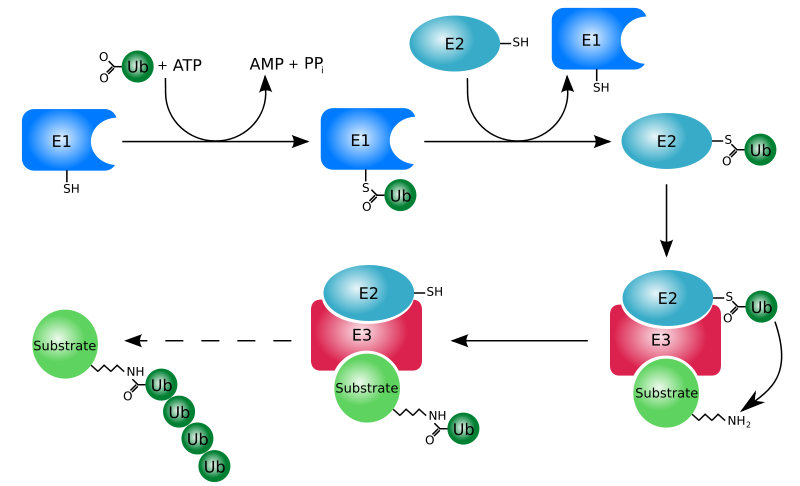
Proces označavanja nekog proteina ubikvitinom (ubikvitinacija) se sastoji od serije koraka:
- Aktivacija ubikvitina: Ubikvitin aktivira E1 ubikvitin-aktivirajući enzim u procesu koji zahtijeva ATP kao izvor energije. Prvi korak uključuje proizvodnju ubikvitin-adenilata. Druga faza je prenos ubikvitina do aktivnog mjesta, cisteina na E1 enzimu, uz otpuštanje AMP-a. Ovaj korak rezultira u tioesterskoj vezi između C-terminalne karboksilne grupe ubikvitina i sulfihidrilnom grupom cisteina na enzimu E1.
- Prenos ubikvitina s E1 do aktivnog mesta cisteina ubikvitin-konjugujućeg enzima E2 putem tran(tio)esterifikacijske reakcije. Genomi sisara imaju 30 - 40 ubikvitin-konjugujućih enzima.
- Završni korak reakcije ubikvitinacije je stvaranje izopeptidne veze između lizina ciljanog proteina i C-terminalnog glicina ubikvitina. Ovaj korak u reakciji zahteva aktiviranje jednog od stotina E3 ubikvitin-protein ligaza (ili skraćeno, ubikvitin ligaza). E3 enzimi deluju kao supstrat za prepoznavanje i imaju sposobnost reagovanja sa E2 i sa supstratom.
U reakciji ubikvitinacije E1 se može povezati sa desetinama E2, koji se mogu povezati sa stotinama E3, hijerarhijski. Ostali ubikvitinu-slični proteini takođe reaguju preko E1-E2-E3 kaskade.
E3 enzimi poseduju jedan od dva moguća domena:
- HECT (engl. Homologous to the E6-AP Carboxyl Terminus) domen
- RING (engl. Really Interesting New Gene) domen
Prenos je moguć na dva načina:
- Direktno s E2, preko katalize RING domena E3 enzima
- Preko E3 enzima, katalizom HECT domena E3. U ovom slučaju, intermedijar kovalentnog E3-ubikvitina se stvara pre prebacivanja na supstrat.
Osim da se samo jedan ubikvitin veže na supstrat, moguće je vezivanje niza molekula ubikvitina, pa tako nastaje poliubikvitinski lanac. Osim toga, neki supstrati su modifikovaani adicijom ubikvitinskih modula s lizinskim ostatkom, u procesu koji se naziva multiubikvitinacija. Kao što je već rečeno, ubikvitin ima 7 lizinskih ostataka. Istorijski, prvobitni tip prepoznatih ubikvitinskih lanaca su bili oni povezani preko lizina 48. Novija istraživanja su otkrila varijetet veza koji uključuje sve moguće lizinske nastavke, a osim toga i linearne lance povezane s N-terminalom ubikvitinskog molekula.[6][7] Radovi izdani 2007. godine pokazuju stvaranje razgranatih lanaca ubikvitina putem različitih veza.[8][9] "Atipični" (nelizinski vezani) lanci ubikvitina se spominju u radu naučnik Ikede i Đikića.[10]
Ubikvitinacijski sistem učestvuje u brojnim ćelijskim procesima, kao što su[11]:
- Razgradnja antigena
- Apoptoza
- Stvaranje organela
- Ćelijski ciklus i deoba
- Transkripciju DNK i njen popravak
- Diferencijacija i razvoj
- Imunski odgovor i upale
- Živčana i mišićna degeneracija
- Morfogeneza živčanih ćelija
- Modulacija receptora na površini ćelija i jonskih kanala
- Odgovor na stres i spoljašnje činioce
- Stvaranje ribozoma
- Virusne infekcije


Najistraživaniji poliubikvitinski lanci, oni povezani preko 48-og molekule lizina, označavaju proteine za razgradnju u procesu koji se zove proteoliza. Barem četiri ubikvitinska molekula moraju biti povezana za lizinske nastavke ciljanog proteina kako bi ga prepoznao 26S-proteasom.[12][13][14][15][16]. Proteasom je kompleksna, bačvasta struktura s dve komore, u kojima se odvija proteoliza. Proteini se brzo razgrađaju u manje peptide (obično duge od 3 - 24 aminokiseline). Ubikvitinski molekuli se odvajaju od proteina pre same razgradnje i recikliraju za dalju upotrebu. Iako je većina proteosomskih supstrata ubikvitinirana, postoje primeri neubikvitiniranih proteina za razgradnju u proteasomu.
Ubikvitin takođe može da označava membranske proteine (na primer, receptore) koje treba odstraniti s membrane, i vrši nekoliko signalnih uloga u ćeliji. Molekuli na ćelijskoj membrani označeni s ubikvitinom su često monoubikvitinizirani, i ove modifikacije često označavaju protein za razgradnju u lizosomu.
Neki genetički poremećaji često povezani s ubikvitinom su:
- Gen čija disfunkcija uzrokuje Angelmanov sindrom, UBE3A, nosi na sebi kod za ubikvitin ligazu (E3) enzmin pod nazivom E6-AP.
- Gen poremećen Von Hipel-Lindauvim sindromom kodira ubikvitin E3 ligazu pod nazivom VHL tumorski supresor ili VHL gen.
- Gen poremećen Lidlovim sindromom dovodi do disregulacije epitelnog Na+ kanala (ENaC) i uzrokuje hipertenziju.
- Osam od trinaest identifikovanih gena čija disfunkcija uzrokuje Fanconijevu anemiju kodira proteina koji formira veliki ubikvitin ligaza (E3) kompleks.
- Mutacije Cullin7 E3 ubikvitin ligaze su povezane s 3-M sindromom, recesivnim poremećajem koji rezultira u retardaciji.[17]
Antitijela za ubikvitin se koriste u histologiji kako bi se identifikovale abnormalne nakupine proteina unutar ćelije, koje su markeri bolesti. Ove nakupine se nazivaju inkluzijska telešca. Primeri za takve nenormalne inkluzije su:
- Neufobrilarna klupka u Alzheimerovoj bolesti
- Levijeva telešca u Parkinskonovoj bolesti
- Pickova telešca u Pickovoj bolesti
- Inkluzije u bolesti motornih neurona i Huntingtonovoj bolesti
- Malorijeva telešca u alkoholnoj bolesti jetre
- Rosenthalska vlakna u astrocitima
- ↑ Kerscher O, Felberbaum R, Hochstrasser M (2006). „Modification of proteins by ubiquitin and ubiquitin-like proteins”. Annual Review of Cell and Developmental Biology 22: 159–80. DOI:10.1146/annurev.cellbio.22.010605.093503. PMID 16753028. Pristupljeno 2011-04-07.[mrtav link]
- ↑ Pickart CM (2001). „Mechanisms underlying ubiquitination”. Annual Review of Biochemistry 70: 503–33. DOI:10.1146/annurev.biochem.70.1.503. PMID 11395416. Pristupljeno 2011-04-07.[mrtav link]
- ↑ „The Nobel Prize in Chemistry 2004”. Nobelprize.org. Pristupljeno 2010-10-16.
- ↑ „The Nobel Prize in Chemistry 2004: Popular Information”. Nobelprize.org. Arhivirano iz originala na datum 2010-10-15. Pristupljeno 2010-10-16.
- ↑ „Official website of Nobel Prize Commitee, list of 2004 winners”. Pristupljeno 2008-04-30.
- ↑ Xu Ping, Junmin Peng (2008-05-01). „Characterization of polyubiquitin chain structure by middle-down mass spectrometry”. Analytical Chemistry 80 (9): 3438–44. DOI:10.1021/ac800016w.
- ↑ Peng Junmin, Daniel Schwartz, Joshua E Elias, Carson C Thoreen, Dongmei Cheng, Gerald Marsischky, Jeroen Roelofs, Daniel Finley, Steven P Gygi (2003). „A proteomics approach to understanding protein ubiquitination”. Nature Biotechnology 21 (8): 921–6. DOI:10.1038/nbt849.
- ↑ Kirisako Takayoshi, Kiyoko Kamei, Shigeo Murata, Michiko Kato, Hiromi Fukumoto, Masato Kanie, Soichi Sano, Fuminori Tokunaga, Keiji Tanaka, Kazuhiro Iwai (2006-10-18). „A ubiquitin ligase complex assembles linear polyubiquitin chains”. The EMBO Journal 25 (20): 4877–87. DOI:10.1038/sj.emboj.7601360.
- ↑ Kim Hyoung Tae, Kwang Pyo Kim, Fernando Lledias, Alexei F Kisselev, K Matthew Scaglione, Dorota Skowyra, Steven P Gygi, Alfred L Goldberg (2007-06-15). „Certain pairs of ubiquitin-conjugating enzymes (E2s) and ubiquitin-protein ligases (E3s) synthesize nondegradable forked ubiquitin chains containing all possible isopeptide linkages”. The Journal of Biological Chemistry 282 (24): 17375–86. DOI:10.1074/jbc.M609659200.
- ↑ Ikeda Fumiyo, Ivan Dikic (2008). „Atypical ubiquitin chains: new molecular signals. 'Protein Modifications: Beyond the Usual Suspects' review series”. EMBO Reports 9 (6): 536–42. DOI:10.1038/embor.2008.93.
- ↑ „Ubiquitin Proteasome Pathway Overview”. Arhivirano iz originala na datum 2008-03-30. Pristupljeno 2008-04-30.
- ↑ Komander D (2009). „The emerging complexity of protein ubiquitination”. Biochem. Soc. Trans. 37 (Pt 5): 937–53. DOI:10.1042/BST0370937. PMID 19754430.
- ↑ Ikeda F, Dikic I (2008). „Atypical ubiquitin chains: new molecular signals. 'Protein Modifications: Beyond the Usual Suspects' review series”. EMBO Rep. 9 (6): 536–42. DOI:10.1038/embor.2008.93. PMC 2427391. PMID 18516089.
- ↑ Xu P, Peng J (2008). „Characterization of polyubiquitin chain structure by middle-down mass spectrometry”. Anal. Chem. 80 (9): 3438–44. DOI:10.1021/ac800016w. PMC 2663523. PMID 18351785.
- ↑ Peng J, Schwartz D, Elias JE, Thoreen CC, Cheng D, Marsischky G, Roelofs J, Finley D, Gygi SP (2003). „A proteomics approach to understanding protein ubiquitination”. Nat. Biotechnol. 21 (8): 921–6. DOI:10.1038/nbt849. PMID 12872131.
- ↑ Kirisako T, Kamei K, Murata S, Kato M, Fukumoto H, Kanie M, Sano S, Tokunaga F, Tanaka K, Iwai K (2006). „A ubiquitin ligase complex assembles linear polyubiquitin chains”. EMBO J. 25 (20): 4877–87. DOI:10.1038/sj.emboj.7601360. PMC 1618115. PMID 17006537.
- ↑ Huber Céline, Dora Dias-Santagata, Anna Glaser, James O'Sullivan, Raja Brauner, Kenneth Wu, Xinsong Xu, Kerra Pearce, Rong Wang, Maria Luisa Giovannucci Uzielli, Nathalie Dagoneau, Wassim Chemaitilly, Andrea Superti-Furga, Heloisa Dos Santos, André Mégarbané, Gilles Morin, Gabriele Gillessen-Kaesbach, Raoul Hennekam, Ineke Van der Burgt, Graeme C M Black, Peter E Clayton, Andrew Read, Martine Le Merrer, Peter J Scambler, Arnold Munnich, Zhen-Qiang Pan, Robin Winter, Valérie Cormier-Daire (2005). „Identification of mutations in CUL7 in 3-M syndrome”. Nature Genetics 37 (10): 1119–24. DOI:10.1038/ng1628.
- MeSH Ubiquitin
- Angelmanov sindrom
- OMIM
- UniProt
- UbiPred Arhivirano 2011-07-17 na Wayback Machine-u
- Ubikvitin Arhivirano 2007-03-15 na Wayback Machine-u
- 7.340 Ubikvitinacija