Arabidopsis thaliana
| Arabidopsis thaliana | |
|---|---|

| |
| Arabidopsis thaliana, inflorescență tulpinală | |
| Clasificare științifică | |
| Regn: | Plantae |
| Diviziune: | Angiospermae |
| Clasă: | Eudicotyledoneae |
| Subclasă: | Rosidae |
| Ordin: | Brassicales |
| Familie: | Brassicaceae |
| Gen: | Arabidopsis |
| Nume binomial | |
| Arabidopsis thaliana (L.), Heynh. | |

| |
| Răsândirea geografică a Arabidopsis thaliana.
Zone în care A. thaliana este nativă Zone în care A. thaliana este aclimatizată Zone în care A. thaliana lipsește | |
| Sinonime | |
| Modifică text |
|
Arabidopsis thaliana (denumită popular gâscariță)[2] este o plantă de talie mică din familia Brassicaceae, originară din Eurasia și Africa.[3][4][5][6][7][8] Este întâlnită frecvent de-a lungul acostamentelor drumurilor, frecvent în zona de câmpie, fiind considerată a fi o buruiană.
Este o specie de plantă anuală cu un ciclu de viață relativ scurt, fiind un organism model utilizat foarte frecvent în studiile de biologia și genetica plantelor. Deși este un organism eucariot multicelular complex, A. thaliana are un genom relativ mic, format din aproximativ 135 de perechi de megabaze.[9] A fost prima plantă al cărei genom a fost secvențiat și este un instrument important pentru înțelegerea biologiei moleculare a multor trăsături ale plantelor, inclusiv dezvoltarea florilor, frunzelor și detectarea luminii, precum și în studiile de genetică și genomică.[3][10][11][12]
Descriere
[modificare | modificare sursă]
Arabidopsis thaliana este o plantă anuală (rareori bienală), care atinge de obicei o înălțime de până la 20-25 cm.[7] Frunzele speciei formează o rozetă la bază, însă prezintă și câteva frunze pe tulpina floriferă. Frunzele bazale sunt de culoare verde până la ușor violacee, au o lungime de 1,5-5 cm și o lățime de 2-10 mm, cu o margine întreagă sau grosier zimțată. Frunzele tulpinale sunt mai mici și lipsite de peduncul, de obicei cu marginea întreagă. Frunzele sunt acoperite cu peri mici, unicelulari, denumiți tricomi. Florile au un diametru de 3 mm și sunt dispuse în inflorescențe de tip corimb, astfel încât structura lor este cea tipică pentru familia Brassicaceae. Fructul este o silicvă de 5-20 mm lungime, care conține 20-30 de semințe.[13][14][15][16] Rădăcinile au o structură simplă, cu o singură rădăcină primară care crește pe verticală în jos, producând ulterior rădăcini laterale mai mici. Aceste rădăcini formează interacțiuni cu bacteriile din rizosferă, cum ar fi Bacillus megaterium.[17]
Întregul ciclu de viață al speciei se încheie în șase săptămâni. Tulpina centrală pe care se dezvoltă florile crește după aproximativ 3 săptămâni, iar florile se autopolenizează în mod natural. În laborator, A. thaliana este uzual cultivată în plăci Petri, în ghivece sau în cultură hidroponică, sub lumini fluorescente sau într-o seră.[18]
-
Inflorescență
-
Aspectul frunzelor tulpinale
-
Aspectul frunzelor bazale
-
Fruct




Taxonomie
[modificare | modificare sursă]
Specia a fost descrisă pentru prima dată în 1577, fiind identificată în Munții Harz de către Johannes Thal (1542-1583), un medic din Nordhausen, Thüringen, Germania. Aceasta a denumit specia Pilosella siliquosa. În 1753, Carl Linnaeus a redenumit planta Arabis thaliana, în onoarea lui Thal (în Species plantarum). În 1842, botanistul german Gustav Heynhold a creat genul Arabidopsis și a clasificat planta în acest gen (în Flora von Sachsen). Denumirea generică de Arabidopsis provine din limba greacă, însemnând „asemănătoare cu Arabis” (genul în care Linnaeus o plasase inițial).
Mii de accesiuni naturale consangvinizate de A. thaliana au fost colectate din întreaga sa zonă de răspândire.[19] Acestea prezintă o variație genetică și fenotipică considerabilă, care poate fi utilizată pentru a studia adaptarea acestei specii la diferite medii.[19]
În limba română, denumirea populară a speciei este gâscariță,[2] deși aceasta poate face referire și la speciile endemice din genul Arabis, precum Arabis alpina și A. hirsuta.[20] Printre alte sinonime ale speciei se numără și: Arabis arcuata, Arabis crantziana, Cardamine pusilla, etc.
Habitat și răspândire
[modificare | modificare sursă]Gâscarița este originară din Europa, Asia și Africa, iar distribuția sa geografică este continuă de la Marea Mediterană până în Scandinavia și din Spania până în Grecia.[21] De asemenea, este nativă în ecosistemele alpine tropicale din Africa și, probabil, din Africa de Sud.[22][23] A fost introdusă și naturalizată în întreaga lume,[24] inclusiv în America de Nord în jurul secolului al XVII-lea.[25]
A. thaliana crește cu ușurință și adesea colonizează soluri stâncoase, nisipoase și calcaroase. Este o specie cu răspândire frecventă în câmpie și zona colinară, fiind xeromezofilă.[26] În general, este considerată o buruiană, datorită răspândirii sale pe scară largă pe câmpurile agricole, pe marginea drumurilor, pe liniile de cale ferată, pe terenurile virane și în alte habitate perturbate,[24][27] dar, datorită capacității sale competitive limitate și a dimensiunilor mici, nu este clasificată ca buruiană dăunătoare.[28] În mod analog cu majoritatea speciilor de Brassicaceae, A. thaliana este comestibilă pentru om, și poate fi consumată în salate sau gătită, totuși nu are o utilizare pe scară largă ca legumă.[29]
Utilizare ca organism model
[modificare | modificare sursă]
Botaniștii și biologii au început să cerceteze specia A. thaliana la începutul anilor 1900, iar prima descriere sistematică a mutanților a fost făcută în jurul anului 1945.[30] A. thaliana este în prezent utilizată pe scară largă pentru diverse studii de știința plantelor, inclusiv a geneticii, evoluției, geneticii populațiilor și dezvoltării plantelor.[31][32][33] Deși A. thaliana ca plantă are o importanță directă redusă pentru agricultură, utilizarea sa ca organism model a revoluționat înțelegerea biologiei genetice, celulare și moleculare a plantelor cu flori.
Primul mutant de A. thaliana a fost documentat în 1873 de către Alexander Braun, acesta descriind un fenotip de floare dublă (gena mutantă a fost probabil gena Agamous, clonată și caracterizată în 1990).[34] Cu toate acestea, Friedrich Laibach (care a publicat numărul cromozomilor în 1907), nu a propus A. thaliana ca organism model până în anul 1943.[35] Studenta sa, Erna Reinholz, a susținut teza de doctorat despre A. thaliana în 1945, descriind prima colecție de mutanți de A. thaliana pe care au generat-o cu ajutorul mutagenezei cu raze X. Laibach și-a continuat contribuțiile importante la cercetarea A. thaliana prin colectarea unui număr mare de accesiuni (adesea denumite în mod discutabil „ecotipuri”). Cu ajutorul lui Albert Kranz, acestea au fost organizate într-o colecție mare de 750 de accesiuni naturale de A. thaliana din întreaga lume.
În anii 1950 și 1960, John Langridge și George Rédei au jucat un rol important în stabilirea A. thaliana ca organism util pentru experimentele biologice de laborator. Rédei a redactat mai multe recenzii științifice care au contribuit la introducerea modelului în comunitatea științifică. Începuturile comunității de cercetare care au avut în prim-plan A. thaliana se bazează pe un buletin informativ denumit Arabidopsis Information Service, care a fost înființat în 1964.[36] Prima conferință internațională privind Arabidopsis a avut loc în 1965, la Göttingen, Germania.
În anii 1980, A. thaliana a început să fie utilizată pe scară largă în laboratoarele de cercetare în domeniul plantelor din întreaga lume. A fost unul dintre mai mulți candidați, printre care se numărau porumbul, petunia și tutunul.[35] Ultimele două erau atractive, deoarece erau ușor de transformat cu ajutorul tehnologiilor actuale de atunci, în timp ce porumbul era un model genetic bine stabilit pentru biologia vegetală. Anul în care A. thaliana a devenit plantă model a fost 1986, deoarece atunci au fost descrise transformarea mediată de ADN-T și prima genă clonată de A. thaliana.[37][38]
Studii de genomică
[modificare | modificare sursă]
Datorită dimensiunii reduse a genomului său și pentru că specia este diploidă, Arabidopsis thaliana este utilă pentru cartografierea și secvențierea genetică. Având aproximativ 157 de perechi de megabaze (MBp) și cinci cromozomi,[41] A. thaliana are unul dintre cele mai mici genomuri dintre speciile vegetale. O perioadă îndelungată s-a crezut că are cel mai mic genom dintre toate plantele cu flori, dar în prezent se cunoaște faptul că cel mai mic genom aparține speciilor din genul Genlisea, ordinul Lamiales, în care specia Genlisea tuberosa, o plantă carnivoră, prezintă p dimensiune a genomului de aproximativ 61 Mbp.[42] A fost primul genom vegetal care a fost secvențiat, finalizat în 2000 de către Inițiativa Genomului Arabidopsis.[43] Cea mai recentă versiune a genomului A. thaliana este menținută de către Arabidopsis Information Resource.[44]
Genomul speciei codifică aproximativ 27.600 de gene codificatoare de proteine și aproximativ 6.500 de gene non-codificatoare.[45] Cu toate acestea, baza de date Uniprot enumeră 39.342 de proteine în cadrul proteomului de referință al Arabidopsis.[46] Dintre cele 27.600 de gene codificatoare de proteine, 25.402 (91,8%) sunt în prezent adnotate ca fiind „semnificative”,[47] deși o mare parte din aceste proteine este probabil doar slab înțeleasă și cunoscută doar în termeni generali (spre exemplu, ca „proteină de legare a ADN-ului fără specificitate cunoscută”). Uniprot enumeră mai mult de 3.000 de proteine ca fiind „necaracterizate” ca parte a proteomului de referință.
Plastomul sau genomul plastidian al speciei A. thaliana este reprezentat de o moleculă de ADN cu o lungime de 154.478 de perechi de baze,[39] o dimensiune întâlnită de obicei la majoritatea plantelor cu flori. Acesta cuprinde 136 de gene care codifică proteine ribozomale din subunitatea mică (rps, în galben, vezi figura), proteine ribozomale din subunitatea mare (rpl, portocaliu), proteine ipotetice open reading frame ale cloroplastului (ycf, lămâie), proteine implicate în reacții fotosintetice (verde) sau cu alte funcții (roșu), ARN-uri ribozomale (rrn, albastru) și ARN-uri de transfer (trn, negru).[40]
Genomul mitocondrial al A. thaliana are o lungime de 367.808 perechi de baze și conține 57 de gene.[48] Există multe regiuni repetate în genomul mitocondrial al Arabidopsis. Cele mai mari repetări se recombină în mod regulat și izomerizează genomul.[49] La fel ca majoritatea genomurilor mitocondriale ale plantelor, genomul mitocondrial al Arabidopsis există ca un aranjament complex de molecule ramificate și liniare suprapuse in vivo.[50]
Studii de genetică
[modificare | modificare sursă]
Transformarea genetică a speciei A. thaliana este o procedură de rutină, și uzual se utilizează Agrobacterium tumefaciens pentru a transfera ADN în genomul plantei. Protocolul actual, denumit „floral dip”, implică simpla scufundare a florilor într-o soluție care conține Agrobacterium purtător al unei plasmide de interes și un detergent.[51][52] Această metodă evită necesitatea culturii de țesuturi sau a regenerării plantelor.
Colecțiile de gene „knockout” (gene blocate) de la A. thaliana reprezintă o resursă unică în biologia plantelor, posibilă datorită disponibilității transformărilor de mare capacitate și a finanțării resurselor genomice. Sediul inserțiilor T-DNA a fost determinat pentru peste 300.000 de linii transgenice independente, informațiile și semințele fiind accesibile prin intermediul bazelor de date T-DNA online.[53] Prin intermediul acestor colecții, sunt disponibili mutanți inserționali pentru majoritatea genelor din A. thaliana.
Formarea tricomilor sau perilor este inițiată de către proteina GLABROUS1. Gene knockout ale acesteia produc plante lipsite de peri, denumite glabre. Acest fenotip a fost deja utilizat în experimentele de editare a genelor, și poate fi de interes pentru a fi utilizat ca marker vizual în cercetare pentru a îmbunătăți metodele de editare genetică, precum CRISPR/Cas9.[54][55]
Alte studii
[modificare | modificare sursă]Dimensiunile reduse ale plantei și ciclul de viață rapid al acesteia sunt aspecte avantajoase pentru cercetare. Din moment ce planta a evoluat ca specie efemeră de primăvară, a fost folosită pentru a dezvolta mai multe soiuri de laborator care au nevoie de aproximativ 6 săptămâni de la germinare până la etapa de producție a semințelor mature. Dimensiunea mică a plantei este convenabilă pentru cultivarea într-un spațiu mic și produce un număr ridicat de semințe. În plus, natura autogamă a acestei plante ajută la experimentele genetice. De asemenea, deoarece o plantă individuală poate produce câteva mii de semințe, fiecare dintre criteriile de mai sus conduce la utilizarea A. thaliana ca organism model în genetică.
A. thaliana este un model de studiu pentru proteinele SNARE din plante. Ca urmare a unor studii s-a arătat faptul că proteinele SNARE sunt implicate în transportul vezicular, iar cele de A. thaliana (denumite AtVTI1a) sunt cel mai probabil esențiale pentru transportul dintre aparatul Golgi și vacuole. Totuși, acest subiect este încă neclarificat, iar rolul proteinelor SNARE vegetale în transportul intracelular este neconcludent.[56]
ADN-ul plantelor este vulnerabil la lumina ultravioletă, iar mecanismele de reparare a ADN-ului au evoluat pentru a evita sau repara leziunile genomului cauzate de aceste radiații. Kaiser et al. au arătat faptul că la A. thaliana dimerii de ciclobutan-pirimidină (CPD) induși de lumina UV pot fi reparați prin exprimarea fotoliazei CPD.[57]
Pe 12 mai 2022, NASA a anunțat că specimene de A. thaliana au germinat și au fost cultivate cu succes în mostre de regolit lunar. Deși plantele au germinat cu succes și au crescut în răsaduri, acestea nu au fost la fel de robuste ca specimenele care au fost cultivate în cenușă vulcanică ca grup de control, deși experimentele au constatat, de asemenea, o anumită variație în plantele cultivate în regolit în funcție de locația din care au fost prelevate probele, deoarece A. thaliana cultivată în regolit recoltat în timpul Apollo 12 & Apollo 17 a fost mai robustă decât cele cultivate în probele prelevate în timpul Apollo 11.[58] În prezent, A. thaliana este studiată pe Stația Spațială Internațională de către Agenția Spațială Europeană, cu scopul de a identifica aspecte legate de creșterea și reproducerea plantelor în condiții de microgravitație.[59][60]
Au fost descrise chiar și dispozitive de tipul „plant-on-a-chip”, în care țesuturile de A. thaliana pot fi cultivate în condiții semi-in vitro'. Utilizarea acestor dispozitive poate contribui la înțelegerea mecanismului de reproducere sexuală la A. thaliana.[61]
Dezvoltare
[modificare | modificare sursă]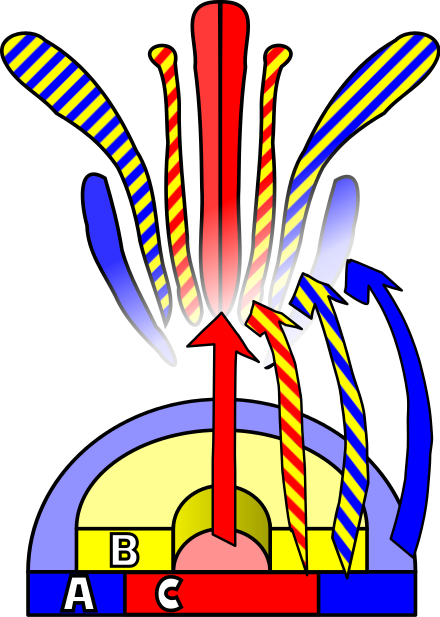
A. thaliana a fost studiată pe scară largă ca model pentru dezvoltarea florilor. Floarea în curs de dezvoltare prezintă patru organe de bază - sepale, petale, stamine și carpele (care vor forma pistilul). Aceste organe sunt dispuse într-o serie de spirale, cu patru sepale pe spirala exterioară, urmate de patru petale în interiorul acesteia, șase stamine și o regiune centrală a carpelelor. Mutațiile homeotice la A. thaliana au ca rezultat transformarea unui organ în altul - în cazul mutației agame, de exemplu, staminele devin petale, iar carpelele sunt înlocuite cu o nouă floare, rezultând un model repetat în mod recursiv de tipul sepală-petală-petală.
Observațiile privind mutațiile homeotice au dus la formularea modelului ABC de dezvoltare florlă, de către E. Coen și E. Meyerowitz.[62] Conform acestui model, genele de identitate a organelor florale sunt împărțite în trei clase: clasa A (care afectează sepalele și petalele), clasa B (care afectează petalele și staminele) și respectiv clasa C (care afectează staminele și carpelele). Aceste gene codifică factorii de transcripție care se combină pentru a determina specificarea țesuturilor în regiunile respective în timpul dezvoltării florale. Deși a fost dezvoltat prin studierea florilor de A. thaliana, acest model se poate aplica, în general, și altor plante cu flori.[63]
Studiile întreprinse asupra A. thaliana au oferit informații considerabile în ceea ce privește genetica morfogenezei frunzelor, în special la plantele dicotiledonate. O mare parte a înțelegerii a provenit din analiza mutanților în etape de dezvoltare a frunzelor, dintre care unii au fost identificați în anii 1960, dar nu au fost analizați cu ajutorul tehnicilor genetice și moleculare până la mijlocul anilor 1990. Frunzele de A. thaliana se pretează bine la studiile privind dezvoltarea frunzelor, deoarece sunt relativ simple și stabile.[64][65] Utilizând A. thaliana, au fost identificate etapele specifice morfogenezei frunzei, fiind trei principale: inițierea unui primordiu foliar, stabilirea dorsiventralității, și dezvoltarea meristemului marginal. Stabilirea dorsiventralității frunzei este extrem de importantă, deoarece din punct de vedere morfologic și fiziologic există diferențe între fața dorsală (adaxială) și cea ventrală (abaxială) a frunzei.[66]
Fiziologie
[modificare | modificare sursă]A. thaliana a fost utilizată pe scară largă pentru studierea bazei genetice a fototropismului, a alinierii cloroplastelor și a deschiderii stomatelor, precum și a altor procese influențate de lumina albastră.[67] Aceste fenomene răspund la lumina albastră, care este percepută de receptorii de lumină ai fototropinei. Arabidopsis a fost, de asemenea, importantă în înțelegerea funcțiilor unui alt receptor al luminii albastre, criptocromul, care este deosebit de important pentru antrenarea luminii pentru a controla ritmurile circadiene ale plantelor.[68] Atunci când debutul întunericului este neobișnuit de devreme, A. thaliana își reduce metabolismul amidonului cu o cantitate care necesită efectiv o diviziune.[69]
Răspunsurile la lumină au fost determinate chiar și în rădăcini, despre care se credea anterior că sunt în mare măsură insensibile la lumină. În timp ce răspunsul gravitropic al rădăcinilor de A. thaliana este răspunsul lor tropic predominant, exemplarele tratate cu mutageni și selectate pentru absența acțiunii gravitropice au prezentat un răspuns fototropic negativ la lumina albastră sau albă și un răspuns pozitiv la lumina roșie, ceea ce indică faptul că rădăcinile prezintă, de asemenea, fototropism pozitiv.[70]
În anul 1990, Janet Braam și Ronald W. Davis au observat faptul că A. thaliana prezintă tigmomorfogeneză ca răspuns la vânt, ploaie și atingere.[71] S-a identificat faptul că cel puțin patru gene induse de atingere există la această specie și sunt reglate de asemenea stimuli.[71] În 2002, Massimo Pigliucci a observat faptul că A. thaliana dezvoltă diverse modele de ramificare ca urmare a expunerii prelungite la vânt, ceea ce este un exemplu de plasticitate fenotipică.[72]
Interacțiunea cu patogeni
[modificare | modificare sursă]Utilizarea speciei A. thaliana a condus la numeroase progrese în cunoașterea modului în care plantele manifestă rezistența la boli. Motivul pentru care cele mai multe plante sunt rezistente la majoritatea agenților patogeni este rezistența în lipsa patogenului, în ideea că nu toți agenții patogeni vor infecta toate plantele. Un exemplu în care A. thaliana a fost utilizată pentru a determina genele responsabile de rezistența la alte plante este Blumeria graminis, agentul cauzal al unei infecții fungice la cereale. Mutanții de A. thaliana au fost dezvoltați cu ajutorul agentului mutagen etil-metansulfonat și au fost examinați pentru a identifica mutanții cu o infecție crescută cu B. graminis. Mutanții cu rate de infecție mai mari sunt denumiți mutanți PEN, datorită capacității lui B. graminis de a pătrunde în A. thaliana pentru a începe procesul de infectare. Genele PEN au fost ulterior cartografiate pentru a identifica genele responsabile de rezistența la B. graminis în lipsa patogenului.[73][74][75]
Note
[modificare | modificare sursă]- ^ Warwick SI, Francis A, Al-Shehbaz IA (). „Brassicaceae species checklist and database”. Species 2000 & ITIS Catalogue of Life (ed. 26). ISSN 2405-8858. Arhivat din original la . Accesat în .
- ^ a b Ciocârlan Vasile (2009). Flora ilustrată a României: Pteridophyta et Spermatophyta. Editura Ceres, București, pag. 533.
- ^ a b „Arabidopsis thaliana”. Germplasm Resources Information Network (GRIN). Agricultural Research Service (ARS), United States Department of Agriculture (USDA). Accesat în .
- ^ Hoffmann, Matthias H. (). „Biogeography of Arabidopsis thaliana (L.) Heynh. (Brassicaceae)”. Journal of Biogeography. 29: 125–134. doi:10.1046/j.1365-2699.2002.00647.x.
- ^ Mitchell-Olds, Thomas (decembrie 2001). „Arabidopsis thaliana and its wild relatives: a model system for ecology and evolution”. Trends in Ecology & Evolution. 16 (12): 693–700. doi:10.1016/s0169-5347(01)02291-1.
- ^ Sharbel, Timothy F.; Haubold, Bernhard; Mitchell-Olds, Thomas (). „Genetic isolation by distance in Arabidopsis thaliana: biogeography and postglacial colonization of Europe”. Molecular Ecology. 9 (12): 2109–2118. doi:10.1046/j.1365-294x.2000.01122.x. PMID 11123622.
- ^ a b Krämer U (martie 2015). „Planting molecular functions in an ecological context with Arabidopsis thaliana”. eLife. 4: –06100. doi:10.7554/eLife.06100
. PMC 4373673
- ^ Durvasula A, Fulgione A, Gutaker RM, Alacakaptan SI, Flood PJ, Neto C, Tsuchimatsu T, Burbano HA, Picó FX, Alonso-Blanco C, Hancock AM (mai 2017). „Arabidopsis thaliana”. Proceedings of the National Academy of Sciences of the United States of America. 114 (20): 5213–5218. doi:10.1073/pnas.1616736114
- ^ „Genome Assembly”. The Arabidopsis Information Resource. Arhivat din original la . Accesat în .
- ^ Hoffmann, Matthias H. (). „Biogeography of Arabidopsis thaliana (L.) Heynh. (Brassicaceae)”. Journal of Biogeography. 29: 125–134. doi:10.1046/j.1365-2699.2002.00647.x.
- ^ Mitchell-Olds, Thomas (decembrie 2001). „Arabidopsis thaliana and its wild relatives: a model system for ecology and evolution”. Trends in Ecology & Evolution. 16 (12): 693–700. doi:10.1016/s0169-5347(01)02291-1.
- ^ „Nifty 50: ARABIDOPSIS -- A PLANT GENOME PROJECT”. www.nsf.gov. Arhivat din original la . Accesat în .
- ^ Flora of NW Europe: Arabidopsis thaliana Arhivat în , la Wayback Machine.
- ^ Blamey, M. & Grey-Wilson, C. (1989). Flora of Britain and Northern Europe. ISBN: 0-340-40170-2
- ^ Flora of Pakistan: Arabidopsis thaliana Arhivat în , la Wayback Machine.
- ^ Flora of China: Arabidopsis thaliana Arhivat în , la Wayback Machine.
- ^ López-Bucio J, Campos-Cuevas JC, Hernández-Calderón E, Velásquez-Becerra C, Farías-Rodríguez R, Macías-Rodríguez LI, Valencia-Cantero E (februarie 2007). „Bacillus megaterium rhizobacteria promote growth and alter root-system architecture through an auxin- and ethylene-independent signaling mechanism in Arabidopsis thaliana”. Molecular Plant-Microbe Interactions. 20 (2): 207–17. doi:10.1094/MPMI-20-2-0207
- ^ Meinke DW, Cherry JM, Dean C, Rounsley SD, Koornneef M (octombrie 1998). „Arabidopsis thaliana: a model plant for genome analysis”. Science. 282 (5389): 662, 679–82. Bibcode:1998Sci...282..662M. CiteSeerX 10.1.1.462.4735
- ^ a b The 1001 Genomes Consortium (iulie 2016). „1,135 Genomes Reveal the Global Pattern of Polymorphism in Arabidopsis thaliana”. Cell. 166 (2): 481–491. doi:10.1016/j.cell.2016.05.063. PMC 4949382
- ^ „Gâscariță” la DEX online Accesat pe 23 martie 2024
- ^ „Arabidopsis thaliana (L.) Heynh”. www.gbif.org (în engleză). Arhivat din original la . Accesat în .
- ^ Hedberg, Olov (). „Afroalpine Vascular Plants: A Taxonomic Revision”. Acta Universitatis Upsaliensis: Symbolae Botanicae Upsalienses. 15 (1): 1–144.
- ^ Fulgione A, Hancock AM (septembrie 2018). „Archaic lineages broaden our view on the history of Arabidopsis thaliana”. The New Phytologist. 219 (4): 1194–1198. doi:10.1111/nph.15244
- ^ a b „Arabidopsis thaliana – Overview”. Encyclopedia of Life. Arhivat din original la . Accesat în .
- ^ Exposito-Alonso M, Becker C, Schuenemann VJ, Reiter E, Setzer C, Slovak R, Brachi B, Hagmann J, Grimm DG, Chen J, Busch W, Bergelson J, Ness RW, Krause J, Burbano HA, Weigel D (februarie 2018). „The rate and potential relevance of new mutations in a colonizing plant lineage”. PLOS Genetics. 14 (2): e1007155. doi:10.1371/journal.pgen.1007155
- ^ lon Sârbu, Nicolae Ștefan, Adrian Oprea (2013). Plante Vasculare din România. Determinator ilustrat de teren. Editura Victor B Victor, București. pag. 466. ISBN 978-606-8149-08-0
- ^ „Arabidopsis thaliana (thale cress)”. Kew Gardens. Arhivat din original la . Accesat în .
- ^ „State and Federal Noxious Weeds List | USDA PLANTS”. plants.sc.egov.usda.gov. Arhivat din original la . Accesat în .
- ^ „IRMNG”. Encyclopedia of Life. Arhivat din original la .
- ^ [1] Arhivat în , la Wayback Machine. TAIR: About Arabidopsis
- ^ Rensink WA, Buell CR (iunie 2004). „Arabidopsis to rice. Applying knowledge from a weed to enhance our understanding of a crop species”. Plant Physiology. 135 (2): 622–9. doi:10.1104/pp.104.040170. PMC 514098
- ^ Coelho SM, Peters AF, Charrier B, Roze D, Destombe C, Valero M, Cock JM (decembrie 2007). „Complex life cycles of multicellular eukaryotes: new approaches based on the use of model organisms” (PDF). Gene. 406 (1–2): 152–70. doi:10.1016/j.gene.2007.07.025. PMID 17870254. Arhivat din original (PDF) la . Accesat în .
- ^ Platt A, Horton M, Huang YS, Li Y, Anastasio AE, Mulyati NW, Agren J, Bossdorf O, Byers D, Donohue K, Dunning M, Holub EB, Hudson A, Le Corre V, Loudet O, Roux F, Warthmann N, Weigel D, Rivero L, Scholl R, Nordborg M, Bergelson J, Borevitz JO (februarie 2010). Novembre J, ed. „The scale of population structure in Arabidopsis thaliana”. PLOS Genetics. 6 (2): e1000843. doi:10.1371/journal.pgen.1000843
- ^ Yanofsky MF, Ma H, Bowman JL, Drews GN, Feldmann KA, Meyerowitz EM (iulie 1990). „The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors”. Nature. 346 (6279): 35–9. Bibcode:1990Natur.346...35Y. doi:10.1038/346035a0. PMID 1973265.
- ^ a b Meyerowitz EM (ianuarie 2001). „Prehistory and history of Arabidopsis research”. Plant Physiology. 125 (1): 15–9. doi:10.1104/pp.125.1.15. PMC 1539315
- ^ „About AIS”. The Arabidopsis Information Resource. . Arhivat din original la . Accesat în .
- ^ Lloyd AM, Barnason AR, Rogers SG, Byrne MC, Fraley RT, Horsch RB (octombrie 1986). „Transformation of Arabidopsis thaliana with Agrobacterium tumefaciens”. Science. 234 (4775): 464–6. Bibcode:1986Sci...234..464L. doi:10.1126/science.234.4775.464. PMID 17792019.
- ^ Chang C, Meyerowitz EM (martie 1986). „Molecular cloning and DNA sequence of the Arabidopsis thaliana alcohol dehydrogenase gene”. Proceedings of the National Academy of Sciences of the United States of America. 83 (5): 1408–12. Bibcode:1986PNAS...83.1408C. doi:10.1073/pnas.83.5.1408
- ^ a b „Arabidopsis thaliana chloroplast, complete genome — NCBI accession number NC_000932.1”. National Center for Biotechnology Information. Arhivat din original la . Accesat în .
- ^ a b Sato S, Nakamura Y, Kaneko T, Asamizu E, Tabata S (). „Complete structure of the chloroplast genome of Arabidopsis thaliana”. DNA Research (în engleză). 6 (5): 283–290. doi:10.1093/dnares/6.5.283
- ^ Bennett MD, Leitch IJ, Price HJ, Johnston JS (aprilie 2003). „Comparisons with Caenorhabditis (approximately 100 Mb) and Drosophila (approximately 175 Mb) using flow cytometry show genome size in Arabidopsis to be approximately 157 Mb, thus approximately 25% larger than the Arabidopsis genome initiative estimate of approximately 125 Mb”. Annals of Botany. 91 (5): 547–57. doi:10.1093/aob/mcg057. PMC 4242247
- ^ Fleischmann A, Michael TP, Rivadavia F, Sousa A, Wang W, Temsch EM, Greilhuber J, Müller KF, Heubl G (decembrie 2014). „Evolution of genome size and chromosome number in the carnivorous plant genus Genlisea (Lentibulariaceae), with a new estimate of the minimum genome size in angiosperms”. Annals of Botany. 114 (8): 1651–63. doi:10.1093/aob/mcu189. PMC 4649684
- ^ The Arabidopsis Genome Initiative (decembrie 2000). „Analysis of the genome sequence of the flowering plant Arabidopsis thaliana”. Nature. 408 (6814): 796–815. Bibcode:2000Natur.408..796T. doi:10.1038/35048692
- ^ „TAIR - Genome Annotation”. Arhivat din original la . Accesat în .
- ^ „Details - Arabidopsis_thaliana - Ensembl Genomes 63”. ensembl.gramene.org (în engleză). Arhivat din original la . Accesat în .
- ^ „Arabidopsis thaliana (Mouse-ear cress)”. www.uniprot.org (în engleză). Arhivat din original la . Accesat în .
- ^ Cheng, Chia-Yi; Krishnakumar, Vivek; Chan, Agnes P.; Thibaud-Nissen, Françoise; Schobel, Seth; Town, Christopher D. (). „Araport11: a complete reannotation of the Arabidopsis thaliana reference genome”. The Plant Journal (în engleză). 89 (4): 789–804. doi:10.1111/tpj.13415
- ^ „Arabidopsis thaliana ecotype Col-0 mitochondrion, complete genome — NCBI accession number BK010421”. National Center for Biotechnology Information. . Arhivat din original la . Accesat în .
- ^ Klein M, Eckert-Ossenkopp U, Schmiedeberg I, Brandt P, Unseld M, Brennicke A, Schuster W (). „Physical mapping of the mitochondrial genome of Arabidopsis thaliana by cosmid and YAC clones”. Plant Journal. 6 (3): 447–455. doi:10.1046/j.1365-313X.1994.06030447.x
- ^ Gualberto JM, Mileshina D, Wallet C, Niazi AK, Weber-Lotfi F, Dietrich A (). „The plant mitochondrial genome: dynamics and maintenance”. Biochimie. 100: 107–120. doi:10.1016/j.biochi.2013.09.016. PMID 24075874.
- ^ Clough SJ, Bent AF (decembrie 1998). „Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana”. The Plant Journal. 16 (6): 735–43. doi:10.1046/j.1365-313x.1998.00343.x. PMID 10069079.
- ^ Zhang X, Henriques R, Lin SS, Niu QW, Chua NH (). „Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method”. Nature Protocols. 1 (2): 641–6. doi:10.1038/nprot.2006.97. PMID 17406292.
- ^ „T-DNA Express: Arabidopsis Gene Mapping Tool”. signal.salk.edu. Arhivat din original la . Accesat în .
- ^ Hahn F, Mantegazza O, Greiner A, Hegemann P, Eisenhut M, Weber AP (). „Arabidopsis thaliana”. Frontiers in Plant Science (în engleză). 8: 39. doi:10.3389/fpls.2017.00039
- ^ Hahn F, Eisenhut M, Mantegazza O, Weber AP (). „Arabidopsis With Cas9-Based Gene Targeting”. Frontiers in Plant Science. 9: 424. doi:10.3389/fpls.2018.00424
- ^ Raikhel, Natasha V. (). „Firmly Planted, Always Moving”. Annual Review of Plant Biology. Annual Reviews. 68 (1): 1–27. doi:10.1146/annurev-arplant-042916-040829
- ^ Kaiser G, Kleiner O, Beisswenger C, Batschauer A. Increased DNA repair in Arabidopsis plants overexpressing CPD photolyase. Planta. 2009 Aug;230(3):505-15. doi: 10.1007/s00425-009-0962-y. Epub 2009 Jun 12. PMID 19521716
- ^ Keeter, Bill (). „Scientists Grow Plants in Lunar Soil”. NASA. Arhivat din original la . Accesat în .
- ^ Link BM, Busse JS, Stankovic B (). „Seed-to-Seed-to-Seed Growth and Development of Arabidopsis in Microgravity”. Astrobiology. 14 (10): 866–875. Bibcode:2014AsBio..14..866L. doi:10.1089/ast.2014.1184. PMC 4201294
- ^ Ferl RJ, Paul AL (aprilie 2010). „Lunar plant biology--a review of the Apollo era”. Astrobiology. 10 (3): 261–74. Bibcode:2010AsBio..10..261F. doi:10.1089/ast.2009.0417. PMID 20446867.
- ^ Yetisen AK, Jiang L, Cooper JR, Qin Y, Palanivelu R, Zohar Y (mai 2011). „A microsystem-based assay for studying pollen tube guidance in plant reproduction”. J. Micromech. Microeng. 25 (5): 054018. Bibcode:2011JMiMi..21e4018Y. doi:10.1088/0960-1317/21/5/054018.
- ^ Coen ES, Meyerowitz EM (septembrie 1991). „The war of the whorls: genetic interactions controlling flower development”. Nature. 353 (6339): 31–7. Bibcode:1991Natur.353...31C. doi:10.1038/353031a0. PMID 1715520.
- ^ John L. Bowman, David R. Smyth, Elliot M. Meyerowitz (November 2012). The ABC model of flower development: then and now. Development. 139 (22): 4095–4098. doi: https://doi.org/10.1242/dev.083972
- ^ Tsukaya H (). „Leaf development”. The Arabidopsis Book. 11: e0163. doi:10.1199/tab.0163. PMC 3711357
- ^ Turner S, Sieburth LE (). „Vascular patterning”. The Arabidopsis Book. 2: e0073. doi:10.1199/tab.0073. PMC 3243335
- ^ Efroni I, Eshed Y, Lifschitz E (aprilie 2010). „Morphogenesis of simple and compound leaves: a critical review”. The Plant Cell. 22 (4): 1019–32. doi:10.1105/tpc.109.073601. PMC 2879760
- ^ Sullivan JA, Deng XW (august 2003). „From seed to seed: the role of photoreceptors in Arabidopsis development”. Developmental Biology. 260 (2): 289–97. doi:10.1016/S0012-1606(03)00212-4
- ^ Más P (). „Circadian clock signaling in Arabidopsis thaliana: from gene expression to physiology and development”. The International Journal of Developmental Biology. 49 (5–6): 491–500. doi:10.1387/ijdb.041968pm
- ^ Scialdone A, Mugford ST, Feike D, Skeffington A, Borrill P, Graf A, Smith AM, Howard M (iunie 2013). „Arabidopsis plants perform arithmetic division to prevent starvation at night”. eLife. 2: e00669. arXiv:1306.5148
- ^ Ruppel NJ, Hangarter RP, Kiss JZ (februarie 2001). „Red-light-induced positive phototropism in Arabidopsis roots”. Planta. 212 (3): 424–30. doi:10.1007/s004250000410. PMID 11289607.
- ^ a b Braam, Janet; Davis, Ronald W. (). „Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis”. Cell (în English). 60 (3): 357–364. doi:10.1016/0092-8674(90)90587-5
- ^ Pigliucci, Massimo (mai 2002). „Touchy and Bushy: Phenotypic Plasticity and Integration in Response to Wind Stimulation inArabidopsis thaliana”. International Journal of Plant Sciences. 163 (3): 399–408. doi:10.1086/339158. ISSN 1058-5893.
- ^ Collins NC, Thordal-Christensen H, Lipka V, Bau S, Kombrink E, Qiu JL, Hückelhoven R, Stein M, Freialdenhoven A, Somerville SC, Schulze-Lefert P (octombrie 2003). „SNARE-protein-mediated disease resistance at the plant cell wall”. Nature. 425 (6961): 973–7. Bibcode:2003Natur.425..973C. doi:10.1038/nature02076. PMID 14586469.
- ^ Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W, Rosahl S, Scheel D, Llorente F, Molina A, Parker J, Somerville S, Schulze-Lefert P (noiembrie 2005). „Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis”. Science. 310 (5751): 1180–3. Bibcode:2005Sci...310.1180L. doi:10.1126/science.1119409. hdl:11858/00-001M-0000-0012-3A32-0
- ^ Stein M, Dittgen J, Sánchez-Rodríguez C, Hou BH, Molina A, Schulze-Lefert P, Lipka V, Somerville S (martie 2006). „Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration”. The Plant Cell. 18 (3): 731–46. doi:10.1105/tpc.105.038372. PMC 1383646
Vezi și
[modificare | modificare sursă]Legături externe
[modificare | modificare sursă]- en Arabidopsis transcriptional regulatory map
- en The Arabidopsis Information Resource (TAIR)
- en Salk Institute Genomic Analysis Laboratory Arhivat în , la Wayback Machine.
- en What Makes Plants Grow? The Arabidopsis genome knows Articol din Genome News Network
- en The Arabidopsis book - A comprehensive review published yearly related to research in Arabidopsis
- en A. thaliana protein abundance
- en The Arabidopsis Information Portal (Araport)
- ro Arabidopsis thaliana