Pteranodon
| Pteranodon Status: Uitgestorven, als fossiel bekend Fossiel voorkomen: Laat-Krijt | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||
| Taxonomische indeling | |||||||||||||||
| |||||||||||||||
| Geslacht | |||||||||||||||
| Pteranodon Marsh, 1876 | |||||||||||||||
| Typesoort | |||||||||||||||
| Pteranodon longiceps Marsh, 1876 | |||||||||||||||
| Afbeeldingen op | |||||||||||||||
| Pteranodon op | |||||||||||||||
| |||||||||||||||
Pteranodon[1][2] is een geslacht van uitgestorven vliegende reptielen, pterosauriërs, behorend tot de groep van de Pterodactyloidea. Het leefde ongeveer 85 miljoen jaar geleden, tijdens het Laat-Krijt, in het gebied van het huidige Noord-Amerika. Het was daarmee een tijdgenoot van grote dinosauriërs, hoewel zelf geen dinosauriër.
Pteranodon werd in 1870 als eerste pterosauriër buiten Europa ontdekt tijdens een avontuurlijke expeditie van de beroemde professor Marsh in het Wilde Westen. Marsh vond eerst alleen stukken bot van de vleugel. Later groeven zijn medewerkers ook schedels op, die lieten zien dat het dier geen tanden had. Daarom gaf Marsh het in 1876 de naam Pteranodon: 'vleugel-zonder-tand'. Daarna werden tot nu toe meer dan twaalfhonderd fossielen van het dier gevonden. Sinds de eerste vondst zijn er vijftien soorten van het geslacht Pteranodon benoemd. Van maar twee daarvan denken we tegenwoordig nog dat het echt aparte soorten zijn: Pteranodon longiceps en de in 1966 benoemde Pteranodon sternbergi. De laatste was misschien de voorouder van de eerste. Sommige wetenschappers zien P. sternbergi echter als een apart geslacht: Geosternbergia. De twee soorten verschillen vooral in de vorm van de schedelkam. Met deze opvallende kam op het achterhoofd liet een dier aan zijn soortgenoten zien hoe oud het was en of het tot de vrouwelijke of mannelijke sekse behoorde.
Pteranodon is de bekendste pterosauriër. Dat komt doordat het met een vleugelspanwijdte van ruim zeven meter tot 1971 het grootste bekende vliegende dier uit de wereldgeschiedenis was. De omvang ervan was zo enorm dat veel geleerden dachten dat het alleen maar korte glijvluchten kon maken en niet eens in staat was goed te zweven. Tegenwoordig begrijpen we dat het lichte maar sterke skelet van Pteranodon voorzien was van krachtige spieren. Hij had een goed uithoudingsvermogen doordat hij, zoals alle pterosauriërs, warmbloedig was en behaard. Zijn vlieghuid had een stelsel van vezels en spiertjes waarmee die steeds op de gewenste spanning gehouden kon worden. Pteranodon kon zo klappend vliegen maar ook met zijn langwerpige vleugels zonder veel inspanning honderden kilometers ver de zee op zweven. Daar viste hij met zijn lange spitse kaken op allerlei zeedieren. Pteranodon is lang niet meer de grootste vliegende soort die we kennen; er zijn pterosauriërs ontdekt die minstens vier keer zo zwaar waren.
Ontdekkingsgeschiedenis en naamgeving
[bewerken | brontekst bewerken]Marsh en Cope
[bewerken | brontekst bewerken]
Het eerste fossiel van Pteranodon, inventarisnummer YPM 1160, werd op 30 november 1870 tijdens een expeditie van Yale College naar Kansas, bij Fort Wallace gevonden door professor Othniel Charles Marsh in de Smoky Hill Chalk van de Niobraraformatie, een mariene kalkafzetting uit het Santonien die doorloopt tot in het vroege Campanien. Het bestond uit een half rechtermiddenhandsbeen van een zeer grote vleugelvinger, die wees op een spanwijdte van zes meter. Ook een tweede middenhandsbeen werd gevonden, YPM 1161. Omdat de kalk in zee afgezet was, zaten er ook resten van vissen tussen. Tanden van een soort daarvan, Xiphactinus audax, werden door Marsh per abuis voor die van de pterosauriër aangezien, omdat hij ervan uitging dat het dier tanden bezeten moest hebben; alle eerdere gevonden pterosauriërs, uit Europa, hadden die immers ook. In 1871 werd de vondst benoemd als Pterodactylus Oweni. In het begin van de negentiende eeuw was het de gewoonte geweest om iedere pterosauriër bij het, eerst benoemde, geslacht Pterodactylus onder te brengen; in 1870 was dat al uit de mode geraakt maar nog wel gebruikelijk voor erg fragmentarische resten waarvan men voorlopig niet precies kon bepalen om wat voor type het eigenlijk ging. De soortaanduiding, indertijd nog vaak met een hoofdletter geschreven, eerde de invloedrijke Britse paleontoloog Richard Owen die zojuist een monografie over de pterosauriër Dimorphodon had gepubliceerd; Marsh deed altijd erg zijn best goede contacten met zijn Europese collegae te onderhouden om zo de status van de nog in haar kinderschoenen staande Amerikaanse paleontologie, waarvan hij zichzelf als de belangrijkste vertegenwoordiger zag, te verhogen.
Tijdens een volgende expeditie in juli en augustus 1871 vond Marsh nog meer resten, waaronder het ontbrekende stuk van het originele middenhandsbeen, YPM 1160, wat de lengte op 412 millimeter bracht. Toen hij die in 1872 wilde beschrijven, werd het hem duidelijk dat de naam Pterodactylus oweni al eerder gebruikt was in 1864. Hij hernoemde daarom P. oweni tot Pterodactylus occidentalis; de soortaanduiding betekent 'de westelijke'. Ook werden twee andere exemplaren benoemd: een grote vleugel die de naam Pterodactylus ingens, 'de ontzaglijke', kreeg en een kleinste vleugel, gevonden op 27 juli, die Pterodactylus velox, 'de snelle', gedoopt werd.

Marsh had een rivaal, Edward Drinker Cope, die indertijd de gewoonte begon te ontwikkelen om steeds wanneer Marsh een vondst gedaan had de aardlagen nog eens zelf te (laten) onderzoeken in de hoop eerder dan deze een naam te kunnen publiceren — of, zoals in dit geval, een vervangende naam. In dit geval mislukte dat net: vijf dagen nadat Marsh's publicatie verscheen, liet Cope op 1 maart 1872 een beschrijving van hetzelfde diertype verschijnen, gebaseerd op vleugelresten die hij in de zomer van 1871 had opgegraven bij Twin Butte Creek. Daarin benoemde hij twee nieuwe soorten van het uit Engeland bekende geslacht Ornithocheirus, een grotere: O. umbrosus en een kleinere: O. harpyia. De keuze voor Ornithocheirus was niet zozeer op een overeenkomst in vorm gebaseerd, hoewel het in beide gevallen om grote soorten met extra lange vleugels ging en Cope door een foute identificatie van een polsbeen dacht dat zijn vondst lange klauwen had als bij de Engelse soort, maar was een poging om een medestander in Engeland te vinden: ook Owen had een rivaal, Harry Govier Seeley, en Ornithocheirus was door de laatste benoemd. Cope maakte echter de fout om de naam fout te spellen als Ornithochirus. Naar huidige normen zou hij zo per ongeluk een apart geslacht geschapen hebben, waarvan de naam niet meer veranderd kan worden. Cope zelf duidde zijn soorten later aan met Pterodactylus. Edward Newton zag het indertijd echter niet zo en verbeterde de spelling in 1888 naar Ornithocheirus. De zaak is echter van weinig belang daar beide soortnamen jongere synoniemen zijn van de soorten van Marsh. Pas na een lang debat wilde Cope, een notoire querulant en ruziemaker, dat in 1875 toegeven maar alleen voor O. harpyia; hij bleef volhouden dat O. umbrosus een aparte soort was.

Op 2 mei 1876 vond een medewerker van Marsh, die zelf in toenemende mate thuisbleef en anderen tussen 1874 en 1880 in Kansas liet opgraven, Samuel Wendell Williston, een eerste, 73 centimeter lange, schedel van de nieuwe soort, YPM 1117, snel gevolgd door een tweede kleinere schedel, YPM 1119. Beiden hadden het verrassende kenmerk dat ze tandeloos waren. Marsh voelde zich hierdoor gerechtigd om dat jaar een nieuw geslacht te benoemen: Pteranodon, afgeleid van het Klassiek Griekse πτερόν (pteron), 'vleugel', en ἀνόδων, anodoon, 'tandeloos' — vanuit ἀν~ (an), 'zonder' en ὀδούς (odous) 'tand'. De drie eerdere soorten Pteranodon occidentalis, P. ingens en P. velox breidde hij uit met een P. longiceps, 'langkaak', voor het grote schedelexemplaar en een P. gracilis, 'lichtgebouwd', voor een nieuw gevonden vleugelfragment. Dat laatste bleek van een erg afwijkende soort te zijn en zou hij al datzelfde jaar hernoemen tot Nyctosaurus. Ook benoemde hij een P. comptus, 'de welgevormde', op basis van twee fragmentarische achterpoten op 17 mei 1875 gevonden door Benjamin Franklin Mudge en P. nanus, 'de dwerg'; de eerste naam wordt tegenwoordig als een nomen dubium beschouwd, de tweede als een mogelijke soort van Nyctosaurus.
Nieuwe geslachten, soorten en ondergeslachten
[bewerken | brontekst bewerken]
In 1893 werd de al erg complexe naamgeving nog ingewikkelder gemaakt doordat Williston de overtuiging kreeg dat een slecht bekende vorm uit Engeland, Ornithostoma, prioriteit zou hebben ten opzichte van Pteranodon. Hij begon dus alle soorten naar dat geslacht te hernoemen. Al in 1901 toonde Felix Plieninger aan dat Williston zich vergist had, maar nog steeds is het mogelijk illustraties te vinden van een Pteranodon-skelet onder de naam van Ornithostoma.
In de twintigste eeuw dook een volgend probleem op: fragmentarische vondsten van grote tandeloze pterosauriërs uit het Krijt werden bij Pteranodon ondergebracht. Later bleken die toe te behoren aan Azhdarchidae. Dat betrof in in 1914 Pteranodon orientalis, die uiteindelijk hernoemd zou worden tot Bogolubovia en in 1928 Pteranodon oregonensis die hernoemd zou worden tot Bennettazhia.
In 1966 werd door John Christian Harksen opnieuw een nieuwe soort benoemd: Pteranodon sternbergi; de soortaanduiding eert George Fryer Sternberg die het fossiel FHSM VP 339, een schedel, in de herfst van 1952 gevonden had bij Bogue. In tegenstelling tot de eerdere namen ging het hierbij echter niet om licht afwijkend materiaal of juist geheel andere soorten, maar om een type dat aan de ene kant sterk leek op wat al van Pteranodon bekend was maar wel een duidelijk afwijkende schedelvorm bezat.

De Niobrarakalksteen is erg productief wat fossielen betreft en ook van Pteranodon zijn vele botten gevonden, in 1910 al 465; in 2004 was dat opgelopen tot boven de 1200 grotere specimina. Behalve uit Kansas zijn die ook gevonden in Wyoming en South Dakota. Hoewel de kwaliteit ervan lang niet altijd even goed is — de vondsten zijn steeds incompleet, soms beschadigd en altijd platgedrukt — maakt dit het geslacht toch een van de best bekende pterosauriërs, samen met Pterodactylus, Rhamphorhynchus en Pterodaustro. Dit verleidde Halsey Wilkinson Miller er in de jaren zeventig toe om een groot aantal subgenera ofwel ondergeslachten in Pteranodon te gaan onderscheiden, zoals Longicepia, Occidentalia en Sternbergia, teneinde wat ordening in het geheel van licht verschillende fossielen te brengen. Miller wilde verder soorten alleen strikt op basis van de schedel onderscheiden, omdat ander materiaal niet eenduidig genoeg was. Van de bestaande soorten erkende hij, afgezien van de in feite tot Nyctosaurus behorende P. bonneri, daarom alleen P. longiceps en P. sternbergi. Hij meende dat dan nog drie typen schedel niet verantwoord waren en daarvoor benoemde hij nog eens drie nieuwe soorten: P. marshi (YPM 2594), P. walkeri en P. eatoni. De laatste was een hernoeming van een schedel, YPM 1179, die Marsh aan P. occidentalis had toegeschreven: de soortaanduiding eert George Francis Eaton die begin twintigste eeuw de eerste gedetailleerde beschrijvingen van het geslacht gaf. P. walkeri was een grote schedel, FHSM VP 221, met kleine kam; de soortaanduiding eert de oorspronkelijke medebeschrijver Myrl Walker.
Het gevolg van dit alles was dat de verwarring alleen maar toenam, vooral toen amateurpaleontoloog George Olshevsky Millers methode tot de uiterste consequentie doorvoerde en alle eerdere soorten bij een van Millers subgenera onderbracht. Deze namen worden tegenwoordig niet meer gebruikt en dat gold lange tijd ook voor het aparte geslacht Geosternbergia dat in 1991 door Olshevsky aan P. sternbergi gegeven werd — nadat Miller het in 1978 als ondergeslacht geschapen had nadat zijn subgenus Sternbergia al een bezette naam bleek — de enige echt te onderscheiden vorm.

In 1972 werd door Ikuwo Obata op grond van een onbepaald been van een ledemaat van een pterosauriër op Hokkaido gevonden in een laag uit het Santonien-Campanien een Pteranodon sp.[ecies] benoemd. Het is onwaarschijnlijk dat er werkelijk enig verband met Pteranodon bestaat.
Bennett schept orde uit de chaos
[bewerken | brontekst bewerken]Volgens een studie van Christopher Bennett uit 1994 kan het Amerikaanse materiaal inderdaad alleen onderscheiden worden op basis van schedelvorm maar zouden daar slechts twee typen in vaststelbaar zijn: dat van Pteranodon longiceps en dat van Pteranodon sternbergi. Alle eerder door Marsh beschreven soorten zijn gebaseerd op materiaal dat de schedel mist en zouden dus ongeldig zijn want nomina dubia. Dat zou Pteranodon longiceps de typesoort maken en Pteranodon sternbergi de tweede soort, de enige andere valide binnen Pteranodon. Zou men echter menen dat beide soorten identiek zijn, bijvoorbeeld omdat de schedelvorm een geslachtsverschil zou weerspiegelen, dan wordt de typesoort P. occidentalis, want YPM 1160 is niet op soort te onderscheiden maar wel zeker van Pteranodon afkomstig. Bennetts indeling werd tot 2010 algemeen aanvaard.
In 2010 echter stelde de Braziliaanse paleontoloog Alexander Kellner dat Geosternbergia toch een apart geslacht was. Daarin onderscheidde hij zelfs twee soorten: Geosternbergia sternbergi en de latere Geosternbergia maiseyi, gebaseerd op specimen KUVP 27821. Tegelijkertijd benoemde hij een apart geslacht voor een specimen, UALVP 24238, dat wegens de hogere ouderdom eerst aan P. sternbergi was toegeschreven: Dawndraco. Kellner merkte ook een specimen op, YPM 2473, dat door Miller aan P. marshi was toegewezen, dat nogal afwijkend was en hij daarom toewees aan een nog onbepaalde Pteranodon sp. Kellners interpretatie compliceert de zaak aanzienlijk.
Soorten
[bewerken | brontekst bewerken]Het resultaat van het naamgevingsproces kan worden uitgedrukt in de volgende lijst van soorten:
| Naam | Naamgever | Jaar | Status | Opmerkingen |
|---|---|---|---|---|
| Pterodactylus occidentalis | Marsh | 1872 | Hernoemd als Pteranodon occidentalis | Eerder Pterodactylus oweni Marsh 1871 (bezet door Seeley 1864), YPM 1160 |
| Pterodactylus ingens | Marsh | 1872 | Hernoemd als Pteranodon ingens | YPM 1170 |
| Pterodactylus velox | Marsh | 1872 | Nomen dubium | Hernoemd als Pteranodon velox, YPM 1176 |
| Ornithochirus umbrosus | Cope | 1872 | Nomen dubium | AMNH 1571 |
| Ornithochirus harpyia | Cope | 1872 | Nomen dubium | AMNH 1572 |
| Pterodactylus umbrosus | (Cope) Cope | (1872) 1874 | Eerder Ornithochirus umbrosus | |
| Pteranodon longiceps | Marsh | 1876 | Valide | Typesoort, holotype YPM 1117 |
| Pteranodon ingens | (Marsh) Williston | (1872) 1876 | Nomen dubium | Eerder Pterodactylus ingens |
| Pteranodon occidentalis | Marsh | (1872) 1876 | Nomen dubium | Eerder Pterodactylus occidentalis |
| Pteranodon velox | Marsh | (1872) 1876 | Nomen dubium | Eerder Pterodactylus velox, juveniel specimen |
| Pteranodon gracilis | Marsh | 1876 | Hernoemd als Nyctosaurus gracilis | |
| Pteranodon comptus | Marsh | 1876 | Nomen dubium | YPM 2335 |
| Pteranodon nanus | Marsh | 1876 | Hernoemd als Nyctosaurus nanus | |
| Ornithocheirus umbrosus | (Cope) Newton | (1872) 1888 | Hernoemd als Pteranodon umbrosus | Spellingscorrectie van Ornithochirus umbrosus |
| Ornithocheirus harpyia | (Cope) Newton | (1872) 1888 | Nomen dubium | Spellingscorrectie van Ornithochirus harpyia |
| Pteranodon umbrosus | (Cope) Williston | (1872) 1892 | Nomen dubium | Eerder Ornithochirus umbrosus |
| Ornithostoma ingens | (Marsh) Williston | (1872) 1893 | Synoniem van Pteranodon ingens | Eerder Pteranodon ingens |
| Ornithostoma umbrosum | (Cope) Williston | (1872) 1897 | Synoniem van Pteranodon umbrosus | Eerder Pteranodon umbrosus |
| Pteranodon oregonensis | Gilmore | 1928 | Hernoemd als Bennettazhia oregonensis | |
| Pteranodon sternbergi | Harksen | 1966 | Valide; wellicht eigen geslacht Geosternbergia | Holotype FHSM VP 339 (eerder SMM 5426) |
| Pteranodon marshi | Miller | 1972 | Synoniem van Pteranodon longiceps | YPM 2594 |
| Pteranodon bonneri | Miller | 1972 | Hernoemd als Nyctosaurus bonneri | |
| Pteranodon walkeri | Miller | 1972 | Synoniem van Pteranodon longiceps | FHSM VP 221 |
| Pteranodon (Occidentalia) eatoni | Miller per Miller | 1972, 1972 | Synoniem van Pteranodon sternbergi | YPM 1179 |
| Pteranodon eatoni | (Miller) Miller | (1972) 1972 | Synoniem van Pteranodon sternbergi | Eerder Pteranodon (Occidentalia) eatoni |
| Pteranodon (Longicepia) longiecps [sic] | Marsh per Miller | 1872, 1972 | Synoniem van Pteranodon longiceps | Eerder Pteranodon longiceps |
| Pteranodon (Longicepia) marshi | Miller per Miller | 1972, 1972 | Synoniem van Pteranodon longiceps | Eerder Pteranodon marshi |
| Pteranodon (Sternbergia) sternbergi | Harksen per Miller | 1966, 1972 | Hernoemd als Pteranodon (Geosternbergia) sternbergi | Eerder Pteranodon sternbergi |
| Pteranodon (Sternbergia) walkeri | Miller per Miller | 1972, 1972 | Hernoemd als Pteranodon (Geosternbergia) walkeri | Eerder Pteranodon walkeri |
| Pteranodon (Pteranodon) marshi | Miller per Miller | 1972, 1973 | Synoniem van Pteranodon longiceps | Eerder Pteranodon marshi |
| Pteranodon (Occidentalia) occidentalis | Marsh per Olshevsky | 1872, 1978 | Synoniem van Pteranodon occidentalis | Eerder Pteranodon occidentalis |
| Pteranodon (Longicepia) ingens | Marsh per Olshevsky | 1872, 1978 | Synoniem van Pteranodon ingens | Eerder Pteranodon ingens |
| Pteranodon (Pteranodon) ingens | Marsh per Olshevsky | 1872, 1978 | Synoniem van Pteranodon ingens | Eerder Pteranodon ingens |
| Pteranodon (Geosternbergia) walkeri | Miller per Miller | 1972, 1978 | Synoniem van Pteranodon longiceps | Eerder Pteranodon walkeri |
| Pteranodon (Geosternbergia) sternbergi | Harksen per Miller | 1966, 1978 | Synoniem van Pteranodon sternbergi | Eerder Pteranodon (Sternbergia) sternbergi |
| Pteranodon orientalis | (Bogolubov) Nesov & Yarkov | (1914) 1989 | Hernoemd als Bogolubovia orientalis | Eerder Ornithostoma orientalis |
| Geosternbergia walkeri | (Miller) Miller per Olshevsky | (1972) 1978, 1991 | Synoniem van Pteranodon sternbergi | Eerder Pteranodon (Geosternbergia) walkeri |
| Geosternbergia sternbergi | (Harksen) Miller per Olshevsky | (1966) 1978, 1991 | Synoniem van Pteranodon sternbergi | Eerder Pteranodon (Geosternbergia) sternbergi |
| Geosternbergia maiseyi | Kellner | 2010 | Valide | Holotype KUVP 27821 eerder beschouwd als specimen P. longiceps |
| Dawndraco kazai | Kellner | 2010 | Valide | Holotype UALVP 24238 eerder beschouwd als specimen P. sternbergi |
Beschrijving
[bewerken | brontekst bewerken]
De algemene proporties van Pteranodon zijn zeer kenmerkend. De schedel is zeer lang, de nek tamelijk kort en ook de romp is gedrongen met een korte staart. De vleugels zijn sterk verlengd maar de achterpoten zwak gebouwd.
Schedel
[bewerken | brontekst bewerken]
Bij de schedel zijn de kaken zeer spits en tandeloos met scherpe kaakranden. Bij het levende dier waren de kaken hoogstvermoedelijk nog verlengd met een hoornbek. Ze zijn licht naar boven gebogen. De bovenkaken zijn vijf à tien procent langer dan de onderkaken: als de bek gesloten is, loopt de kromme onderkant van het uitstekende deel van de bovenkaken precies over in de gekromde kaaklijn van de onderkaken. Een studie uit 2017 toonde aan dat de beenwand van de kaken bestond uit twee lagen met tussenliggende trabeculae, beenstutten. Bij groeiende dieren werd er op de buitenste laag aan de buitenkant in een golfbeweging naar voren toe bot afgezet terwijl de dikte constant bleef doordat er gelijktijdig aan de binnenkant bot geresorbeerd werd. Bij jongvolwassen exemplaren leidt dat tot en opvallend streeppatroon dat bij oudere dieren echter verdwenen is.
Ondanks de algemene lichte bouw zijn de schedelopeningen klein, zodat het geheel toch een vrij massieve indruk maakt zonder duidelijke schedelnaden. De processus nasalis, een uitsteeksel van het neusbeen in de fenestra nasoantorbitalis, de voorste schedelopening, is sterk gereduceerd. De relatief kleine oogkassen staan vrij hoog. De snuit is hol zonder echte voorste kam — hoewel de bovenzijde een scherpe rand vormt — en loopt zeer geleidelijk over in een kam die vooraan ontspruit aan de voorhoofdsbeenderen en achteraan zijn basis heeft aan het achterhoofd. Bij P. longiceps is de vol uitgegroeide kam erg lang, slechts drie millimeter dik, smal en naar achteren nog versmallend. De onderkaken zijn over het voorste twee derde deel van hun lengte vergroeid tot een symfyse. Ze hebben onderaan een lage kam die in een punt uitloopt, die de aanhechting zou kunnen zijn geweest van een keelzak. Het tongbeen dat zo'n zak had moeten ondersteunen is echter zwak gevormd en slechts gedeeltelijk verbeend, wat aan de andere kant weer de flexibiliteit ervan verhoogt. Het onderste slaapvenster is spleetvormig en zit boven een vrijwel liggend quadratum dat het bovenste deel van het kaakgewricht vormt. Dat is schroefvormig zodat de onderkaken bij het openen automatisch uiteen gedrongen worden, wat de keelholte vergroot, een aanpassing aan het heel inslikken van een prooi.
Postcrania
[bewerken | brontekst bewerken]
De delen achter de schedel, de postcrania, zijn goed bekend maar slechts zelden met een bepaalde schedel in verband te brengen. Alle wervels zijn sterk gepneumatiseerd, doortrokken van luchtholten, met foramina pneumatica, openingen waardoor vertakkingen van de luchtzakken het holle bot kunnen binnendringen. Het voorkomen van luchtzakken, die een goede zuurstofvoorziening garanderen door steeds verse lucht in één richting door de stijve longen te pompen, is een sterke aanwijzing dat de soort warmbloedig was en voorzien van een haarvacht. Er zijn negen halswervels waarvan de zijdelingse beweging verminderd wordt door exapofysen, uitsteeksels aan de zijkanten. De atlas en de draaier zijn vergroeid. De halswervels hebben vrij hoge doornuitsteeksels. De nek is ongeveer half zo lang als de romp.

Er zijn twaalf ruggenwervels waarvan de voorste zes tot acht zijn samengegroeid tot een notarium; de doornuitsteeksels ervan zijn vergroeid tot een supraneurale plaat; tussen de derde en vierde wervel bevindt zich daarop een gewrichtsfacet voor de articulatie met het schouderblad. Drie tot vier paar ribben zijn aan het notarium vastgegroeid. Schouderblad en het iets langere via naast elkaar liggende gewrichtsvlakken met het borstbeen articulerende ravenbeksbeen zijn tot een geheel vergroeid. Zo ontstaat een gesloten beenring die de krachten die het slaan met de vleugels op de borstkas zou kunnen uitoefenen, afvoert naar notarium en borstbeen. Dit soort structuren is bij alle Pterosauria bekend maar Pteranodon is hierin sterk gespecialiseerd. Het borstbeen is trapeziumvormig, bijna vierkant, en loopt van voren uit in een lange punt, de cristospina, die als aanhechting voor de vliegspieren dient. Dit punt is daartoe ruw van oppervlak. Een echte beenkiel is er niet op het borstbeen, wel een middenrichel van tot vier millimeter hoog.

Het vrij rechte opperarmbeen draagt een grote deltopectorale aanhechtingskam voor de vliegspieren en is daardoor bijlvormig. Om het aanhechtingsvlak nog te vergroten is de kam van onderen naar binnen gedraaid en verbreed. Ook het opperarmbeen is gepneumatiseerd met aan de onderkant van beide uiteinden een groot foramen pneumaticum. Het opperarmbeen neemt richting onderarm in diameter toe en heeft op het einde een driehoekig puntje. De onderarm is een derde langer dan het opperarmbeen. In de onderarm is de ellepijp langer dan het spaakbeen, dat relatief forsgebouwd is. De middenhandsbeenderen zijn anderhalf keer langer dan de onderarm. De drie eerste middenhandsbeenderen zijn gereduceerd, splintervormig, raken de pols niet meer en dragen slechts kleine vingers met zwakke klauwen. De formule voor de vingerkootjes is, inclusief klauwen, 2-3-4-4-x. Alle polsbeenderen zijn versmolten tot een bovenste en onderste rij. Van de pols loopt een lang os pteroides richting nek, als ondersteuning voor een propatagium, een membraan tussen de voorste vleugelrand en de hals. Dit pteroïde is met de pols verbonden via een kogelgewricht. Het vierde middenhandsbeen draagt de vleugelvinger die vier kootjes heeft. Het eerste kootje is langer dan het middenhandsbeen en daarmee het langste bot in de vleugel. Het tweede kootje is juist iets korter; het derde en vierde kootje lopen snel in lengte af. Het vierde kootje eindigt in een vrij scherpe naar binnen gebogen punt. Een dergelijke afschuining vermindert de weerstand bij lage snelheden. Een aantal exemplaren laat geheelde breuken zien in het vierde kootje; dat was kennelijk extra kwetsbaar maar een breuk erin was dus niet fataal. Aan de vleugelvinger is de vlieghuid verbonden. Tot hoever die doorliep: heup, staart, knie of enkel, is omstreden.

Er zijn zes sacrale wervels. Het darmbeen heeft een lang voorste uitsteeksel voor de aanhechting van de beenspieren. Dat ligt naast vier ruggenwervels die vroeger wel bij de sacrale wervels gerekend werden. Het achterste uitsteeksel is bij volwassen exemplaren vergroeid met de echte sacrale wervels, een synsacrum vormend. De staart is kort, ongeveer vijfentwintig centimeter lang, en taps toelopend met minimaal elf staartwervels waarvan de uiterste drie zijn vergroeid tot een enkel beenelement waarvan het uiteinde bestaat uit twee naast elkaar liggende lange smalle staafvormige uitsteeksels en dat de helft van de staartlengte uitmaakt. Deze dienen wellicht als aanhechting voor membranen; volgens Bennett en Padian vermoedelijk die van de vleugels, volgens de meesten voor twee vliezen die van beide enkels tot aan de staart liepen. De staartbasis is echter flexibel en bestaat uit korte wervels, waarvan de voorste bij grotere individuen aan het bekken is vastgegroeid. Deze speciale vorm van de staart is pas sinds de jaren tachtig bekend en eerdere illustraties tonen de staart te kort.
Het dijbeen is maar iets langer dan het opperarmbeen. Het anderhalfmaal langere scheenbeen is vergroeid met een zeer kort rudimentair kuitbeen. De middenvoetsbeenderen zijn zwak gevormd en de voet is klein, korter dan het dijbeen, hoewel langer dan bij andere gespecialiseerde zwevers zoals de Ornithocheiridae. De formule van de teenkootjes is 2-3-4-5-x. Van de vijfde teen is alleen een kort maar dik middenvoetsbeen over. De teenklauwen zijn vrij klein en niet sterk gekromd. Tezamen is de achterpoot iets langer dan de romp.

De vleugelspanwijdte was rond de zevenenhalf meter voor P. longiceps en misschien wel boven de acht meter voor P. sternbergi. Pteranodon was daarmee een van de grootste van de geslachten uit de Pterosauria. Dit zijn echter de maximale afmetingen van gevonden exemplaren; de meeste volwassen individuen bleven veel kleiner. Het grootste exemplaar van P. longiceps is YPM 2833, bestaande uit een eerste vingerkootje van 692 millimeter en een tweede kootje van 545 millimeter, die samen op een spanwijdte van ongeveer zevenhonderdzestig centimeter duiden. Eaton leidde uit de grootste fragmentarische vleugel hem bekend een spanwijdte van 816 centimeter af maar dat wordt tegenwoordig als een overschatting gezien.
Sommige fossielen laten versteende stukken van de weke delen zien. Zo zijn bij specimen KOVP 968 gefossiliseerde gedeeltelijk verbeende pezen van de armspieren zichtbaar.
Over de kleur van de vacht van Pteranodon is niets bekend. In de negentiende eeuw werd die meestal als bruin verondersteld; in het midden van de twintigste eeuw, toen men dacht dat het dier helemaal geen vacht had maar bedekt was met schubben, stelde men zich Pteranodon groen voor, de sterotypische standaardkleur voor reptielen. Nadat in het midden van de jaren tachtig het vogelmodel gebruikt werd om pterosauriërs mee te vergelijken, gaf men een zeebewoner als Pteranodon in illustraties vaak de typische tegenschaduwing van huidige zeevogels: zwart van boven en wit van onderen. Misschien dat de kam een fel kleuraccent droeg als signaalkleur.
Wijze van voortbewegen
[bewerken | brontekst bewerken]Vliegen
[bewerken | brontekst bewerken]
Toen Pteranodon in 1870 ontdekt werd, was het de grootste bekende pterosauriër. In 1958 werd Titanopteryx beschreven, later herdoopt tot Arambourgiania, maar die vondst bestaat slechts uit een enkele halswervel. Pas met de bekendmaking van Quetzalcoatlus in 1975 zou het duidelijk worden welke enorme omvang dergelijke Azhdarchidae konden bereiken. Tot die tijd vertegenwoordigde Pteranodon de vermeende bovenlimiet voor vliegende dieren.
In de late negentiende eeuw fungeerde het dier daarbij als een inspiratiebron voor de luchtvaartpioniers. De mens was nog niet in staat vliegtuigen te bouwen die groter waren dan een speelgoedmodel maar Pteranodon liet, meer dan de huidige vogels, zien dat een 'zwaarder-dan-lucht'-systeem een aanzienlijke grootte kon bereiken. Er was in die jaren, voordat lichte en sterke benzinemotoren werden ontwikkeld, vooral veel belangstelling voor het zweefvliegtuig en ook Pteranodon werd allereerst als een zwever voorgesteld. In 1914 verscheen een studie van het vliegvermogen van dit dier door Ernest Hanbury Hankin en David Meredith Seares Watson waarbij dit aspect sterk op de voorgrond trad. Dit zou voor vele jaren de trend zetten. Omdat pterosauriërs in het midden van de twintigste eeuw als koudbloedig werden gezien, leek veel meer dan een glijvlucht ook niet tot de mogelijkheden te behoren en dacht men dat Hankin en Watson nog optimistisch waren geweest door een goed langdurig zweefvermogen te veronderstellen.

In de vroege jaren zeventig publiceerde Cherrie Bramwell een aantal studies waarin zij een meer kwantitatieve en exacte benadering van het probleem probeerde te geven. Allereerst berekende zij de maximale krachten die het vleugelskelet binnen een zekere veiligheidsmarge kon doorstaan. Dat het opperarmbeen hol is, met een beenwanddikte van nog geen millimeter, resulteerde in een lage schatting van het piekvermogen van de vliegspieren: niet meer dan vijfenzeventig watt bij een pteranodon met een spanwijdte van zeven meter. Bramwell leidde hieruit af dat het dier niet goed tot een klappende vlucht in staat was en compenseerde dit door een zo groot mogelijke vlieghuid te veronderstellen, waarvan de achterrand in een rechte lijn van de vleugelpunten tot de enkels liep. Dat leverde een vleugeloppervlakte op van vijf vierkante meter. Doordat er maar drie kilo aan vliegspieren verondersteld werd, was het geschatte gewicht slechts 16,6 kilogram en de vleugelbelasting aldus extreem laag, minder dan bij echte zweefvliegtuigen. De grote vleugels resulteerden in een hoge luchtweerstand. Bramwell berekende dat het dier zestig watt had moeten leveren om met de vleugels slaand horizontaal te kunnen vliegen bij volledige windstilte. Het zou dan een voorwaartse snelheid van vijfendertig km/u hebben gehad, zo laag dat het nog last zou hebben gehad van een aanzienlijke geïnduceerde weerstand. Restvermogen om op te stijgen of een prooi te vangen was er dan nauwelijks en iedere winderige dag zou het gedwongen hebben op de grond te blijven. Zo bleef voor Pteranodon alleen een levenswijze als pure zwever open. Dat zou ook het schoudergewricht bewijzen waarin zich volgens Bramwell een speciale horizontale groef bevond waarin het opperarmbeen zich zou hebben kunnen vergrendelen in een vaste positie om zonder de vleugels te bewegen rond te zweven. Het dier zou alleen met enige regelmaat hebben kunnen opstijgen door zich van klippen te werpen. Diezelfde klippen zouden dan de stijgwinden hebben moeten leveren om met een prooi weer terug te keren. Die moest dan in een korte vlucht gevangen worden want voor dynamisch zweven, het gebruikmaken van snelheidsverschillen tussen luchtlagen, was de vleugelbelasting te laag; het kon te weinig snelheid winnen in een duikvlucht. De vleugels gedeeltelijk inklappen om hun oppervlakte te verminderen, zou tot een volledig verlies aan dragend vermogen geleid hebben omdat dan de vlieghuid niet meer onder spanning gehouden had kunnen worden. Pteranodon zou, ook als warmbloedigheid aangenomen werd, zo de bovengrens bereikt hebben waarop een vliegend dier nog redelijk kon functioneren en de soort als geheel had zich eigenlijk alleen kunnen handhaven doordat de meeste individuen een stuk kleiner waren dan zeven meter spanwijdte.
Op Bramwells interpretatie kwam al meteen veel kritiek. Er zijn geen aanwijzingen dat zich in het gebied waar Pteranodon leefde klippen bevonden; de kust was gevormd door rivierdelta's. Daarbij zijn de fossielen tot honderden kilometers van de toenmalige kustlijn aangetroffen. Het dier moest dus minstens tot dynamisch zweven in staat zijn geweest. De vermeende groeve in het schoudergewricht is de normale beennaad tussen schouderblad en ravenbeksbeen die bij iedere pterosauriër voorkomt en bij leven bedekt was met kraakbeen en ander weefsel dat een vergrendeling uitsloot.

In de jaren tachtig en negentig werd duidelijk waarin Bramwells analyse fundamentele fouten gemaakt had. De belangrijkste vergissing was het aannemen dat het opperarmbeen volledig hol was. Bramwell had zich al verbaasd dat dit essentiële bot zwakker geconstrueerd was dan de overige vleugelbeenderen. In feite was het gevuld met sponsachtig botweefsel waardoor het minstens driemaal sterker was dan door haar berekend. Bramwell had dit niet direct kunnen waarnemen doordat de fossielen van Pteranadon zo platgedrukt waren; latere vondsten van andere grote pterosauriërs zouden de werkelijke structuur openbaren. Een sterker opperarmbeen maakte veel zwaardere vliegspieren mogelijk en ook een hoger totaalgewicht van ongeveer dertig kilo bij een vleugelspanwijdte van zeven meter. Mark Paul Witton schatte het gewicht in 2008 zelfs op zesendertig kilo bij een vlucht van zes meter. Die spieren zouden een 150 à 200 watt hebben kunnen leveren. Het minimumvermogen om horizontaal te kunnen blijven vliegen, steeg echter vermoedelijk niet boven de door haar berekende zestig watt; het hogere gewicht werd gecompenseerd door een aanzienlijk efficiëntere vleugelvorm met een veel lagere luchtweerstand. James Brower en Kevin Padian namen aan dat de vlieghuid aan de heup bevestigd was. Zelfs indien hij tot aan de enkels reikte, kon de vleugel slank zijn doordat het weefsel niet elastisch was, zoals Bramwell had aangenomen, maar gevormd door een laag van voor naar achteren lopende verdikte richels verbonden door spiertjes. Dat maakte dat de vleugel voldoende stijf kon zijn en toch een bocht kon maken van de elleboog naar de voet. Ook kon door de spiertjes samen te trekken het vleugelprofiel aan de omstandigheden worden aangepast en bleef het dragend vermogen bij het gedeeltelijk inklappen behouden. De veel hogere vleugelbelasting maakt dynamisch zweven mogelijk en ook een hogere kruissnelheid van tussen de vijftig en zestig km/u, voor dit geval het meest efficiënte snelheidsinterval met de laagste combinatie van geïnduceerde en parasitaire weerstand. Pteranodon kwam dus op veel punten overeen met de huidige grote dynamische zwevers, de albatrossen, ook viseters met een grote vleugelslankheid die vaak als interpretatiemodel gebruikt worden. Er zijn echter ook belangrijke verschillen: Pteranodon was relatief toch veel lichter gebouwd — een albatros met een vlucht van zeven meter zou honderd kilo gewogen hebben — maar wel ten opzichte van zijn gewicht veel zwaarder gespierd. Ondanks zijn grotere absolute omvang zou hij dus in staat geweest zijn lange periodes van zweefvlucht te onderbreken door een klappende vlucht. Zeker de kleinere exemplaren konden die langdurig volhouden.

Ook de gedachte dat Pteranodon klippen althans nodig gehad zou hebben om op te stijgen, is verlaten. Die klippen, en de versnellende zwaartekracht die ze konden verschaffen, leken nodig te zijn doordat Pteranodon veel te zwakke benen had om met een sprong van de achterpoten het luchtruim te kiezen, zoals de meeste vogels doen. Zich al klapwiekend van het aardoppervlak verheffen was voor zo'n grote vorm vrijwel uitgesloten. Een kopwind zou het dier als het de vleugels spreidde al snel hebben kunnen wegblazen — bij Bramwells schattingen zou het dier 'zo licht als een vlinder' beladen zijn geweest en zich dus zelfs voor sterke winden hebben moeten verschuilen. Maar als het door een bries opgetild werd, zou het meestal meteen weer op de aarde neergesmeten zijn; alleen bij hoge uitzondering zou een storm het dier hoog genoeg opgenomen hebben dat het daarna in een duikvlucht de minimale vluchtsnelheid ten opzichte van de bewegende lucht had kunnen bereiken. De voorwaartse snelheid ten opzichte van het aardoppervlak is op zich irrelevant voor de vraag of men in de lucht kan blijven. Bramwell had al overwogen of niet de veel krachtiger vleugels zelf voor de afzet gebruikt hadden kunnen worden, maar deze mogelijkheid verworpen, opnieuw vanwege de vermeende zwakte van het opperarmbeen. In het begin van de eenentwintigste eeuw is het door computersimulaties en het onderzoek van Michael Habib duidelijk geworden dat deze vorm van opstijgen voor de meeste pterosauriërs, en vooral de grotere soorten, de gebruikelijke of zelfs enig mogelijke moet zijn geweest. Als de romp op de lange armen leunde, waren die sterk genoeg om in één afzetbeweging tegen de grond het lichaam met vijftig km/u onder een hoek van dertig graden meters de lucht in te slingeren. Zo werd onmiddellijk de meest efficiënte snelheid bereikt en meteen de hoogte gewonnen om de vleugels naar voren en beneden uit te slaan, waarmee de vlucht een feit was. Het ook op de armen staan was daarbij de normale loophouding.
Lopen
[bewerken | brontekst bewerken]
De achterpoten van pterosauriërs tonen een zekere spreidstand, de dijbenen zijn meer zijwaarts gericht. Bij de vleugel konden de opperarmbeenderen niet strak tegen het lichaam ingeklapt worden en in een ingetrokken positie in ieder geval niet goed van voor naar achteren bewegen. Bij het lopen moesten dus ook deze zijwaarts gehouden worden. Aangezien de vrij stijve romp niet gekromd kon worden, waren de eerste pterosauriërs zo beperkt tot een tamelijk langzame stappende gang met het lichaam vrij laag boven de grond. Bij de latere Pterodactyloidea werden de benen korter en de vleugels langer; dat laatste vooral door een enorme verlenging van het middenhandsbeen. Ondanks deze verschillende proporties namen de meeste geleerden aan dat ook de pterodactyloïden de romp laag hielden. Dat kon alleen door ook de middenhandsbeenderen zijwaarts uit te klappen. Dat zou voor een grote soort als Pteranodon met ook nog eens extra lange vleugels, geresulteerd hebben in een uitermate onbeholpen gang waarbij het dier van voren voornamelijk over de bodem schoof en zich van achteren met zijn kromme beentjes wat voortduwde. Men dacht daarom dat het beest de grond zo veel mogelijk meed. Met de korte handklauwen was het daarbij uitgesloten om in een boom te klauteren. Bramwell veronderstelde dat Pteranodon bij voorkeur ondersteboven aan een rotswand hing.


In de jaren tachtig deed Padian het eerdere idee van Seeley herleven dat pterosauriërs tweevoeters waren die zich op de achterpoten voortbewogen en teengangers waren. Die hypothese leek niet onwaarschijnlijk voor kleinere vroege vormen als Dimorphodon, die lange benen en staart hadden. Padian stelde echter dat ook Pteranodon op twee benen liep en zelfs rennend en klapwiekend opsteeg.
De ichnologie, de leer van het onderzoek naar fossiele sporen, heeft echter duidelijk gemaakt dat althans de Pterodactyloidea op vier poten liepen. Dat deden ze echter niet op een onbeholpen wijze. De middenhandsbeenderen werden meer verticaal gehouden zodat de romp een opgerichte houding kreeg. Ook de achterpoten stonden rechter dan eerst werd aangenomen. Wel waren het zoolgangers: de voetzolen stonden dus plat op de grond. Zo kon Pteranodon met een redelijke snelheid voortstappen en misschien was zelfs een draf mogelijk. Gecombineerd met het vermogen om vanuit die loophouding snel op te stijgen was het dier ook niet bijzonder kwetsbaar voor roofdieren, zoals de grote theropoden — hoewel er daarvan uit die tijd toevallig geen uit Noord-Amerika bekend zijn.
Functie van de kam
[bewerken | brontekst bewerken]De schedelkam is een opvallend kenmerk van Pteranodon. Tegenwoordig weten we dat veel pterosauriërs, groot en klein, met kammen op het hoofd waren uitgerust maar in de negentiende eeuw was dat nog onbekend, zodat men een speciale verklaring ging zoeken waarom juist Pteranodon een kam had.
Eaton opperde in 1910 in zijn grote studie over de osteologie van de soort twee mogelijke functies: een aanhechtingspunt voor lange kaakspieren — immers de normale rol van de wandbeenderen — of een contragewicht voor de lange kop, waarbij vooral de luchtweerstand de nodige druk had moeten opleveren want op zich was de dunne kam niet zwaar. Hij had daarbij een voorkeur voor de laatste verklaring want hij kon geen reden bedenken waarom het dier kaakspieren met een groter uithoudingsvermogen (de kracht neemt door de lengte niet toe) nodig gehad moest hebben; het was wel niet om te kauwen met de tandeloze bek die alleen op en neer kon bewegen.
In 1943 stelde Dominik von Kripp dat de kam functioneerde als richtingsroer. Dit werd een populaire theorie want aerodynamisch gezien zou een groot stuurvlak in een positie nog voor de vleugels een grote wendbaarheid moeten hebben opgeleverd.[3] Ross Stein suggereerde in 1975 zelfs dat om dit effect te versterken de kam een verankering vormde voor een membraan dat naar achteren tot de nek of zelfs de rug liep, hoewel dit de verticale beweeglijkheid van de nek sterk gehinderd zou hebben. Het zou zo wel gediend kunnen hebben als een effectieve luchtrem om vlug snelheid te verminderen.
De kam was doorspekt met bloedvaten. Volgens Kellner werden dergelijke kammen daarom gebruikt als radiator om een teveel aan lichaamswarmte uit te stralen. Bennett heeft er echter op gewezen dat al deze verklaringen niet het feit in beschouwing nemen dat de meeste exemplaren maar een vrij kleine kam hebben. De aerodynamische en thermodynamische effecten zouden hoe dan ook beperkt geweest zijn. Hij acht het het waarschijnlijkst dat de kam, wellicht fel gekleurd, diende ter communicatie binnen de soort en leeftijd of geslacht aanduidde.
Seksuele dimorfie
[bewerken | brontekst bewerken]

De overvloed aan fossiel materiaal maakt het mogelijk statistische analyses ervan uit te voeren. Volgens studies van Bennett uit 1991 en 1992 zouden er in de populatie twee grootteklassen aan te wijzen zijn, onafhankelijk van de leeftijd die door de mate van verbening is vast te stellen; een grafiek van de lichaamsgrootte van volwassen exemplaren leverde dus een bimodale verdeling op, met twee pieken. Hij mat dit onder andere aan de doorsnede van het middenhandsbeen: bij de ene groep lag de modus op vijfentwintig, bij de andere op vijfendertig millimeter; die was dus ruim tweemaal zo zwaar met een gemiddelde vlucht van ruwweg vijf meter tegenover zo'n drieënhalve meter voor de groep kleinere individuen. De twee klassen hingen weer samen met verschillen in de vorm van de kam en de breedte van het bekken. Dit vertegenwoordigde ook geen evolutionaire reeks; het verschijnsel deed zich door de hele formatie heen voor, onafhankelijk van de geologische ouderdom. Bennett concludeerde hieruit dat deze verdeling veroorzaakt werd door seksuele dimorfie, een onderscheid in bouw tussen de geslachten. De klasse met de kleinste bouw had de breedste bekkens en de kleinste kam; dit zouden de wijfjes zijn geweest, die immers een breed bekkenkanaal nodig hadden om eieren te leggen. De grotere exemplaren waren dan de mannetjes, die met een hun grote kam indruk zouden hebben gemaakt op de wijfjes, in een proces van seksuele selectie, of op hun mannelijke rivalen als dominantiesignaal. Opmerkelijk is dat, als deze interpretatie correct is, er zich kennelijk tweemaal zoveel wijfjes als mannetjes in de populatie bevonden. Bennett zag dat als een teken dat de soort polygaam was, waarbij de mannetjes een harem probeerden te vormen.
Een probleem is dat door de compressie van de fossiele skeletten de vorm van het bekken lastig is vast te stellen. Ook hebben bij de meeste groepen reptielen juist de mannetjes het breedste bekken. Kellner meende in 2010 dat er te weinig exemplaren met zowel schedel als bekken gevonden waren om een significant verband tussen kamvorm en bekken te kunnen vaststellen. Dit zou vooral een probleem opleveren bij P. sternbergi waarvan veel minder exemplaren bekend zijn.
Pteranodon sternbergi of Geosternbergia
[bewerken | brontekst bewerken]
Volgens de analyse van Bennett zijn er, onafhankelijk van het geslacht, twee morfen, bouwtypen, te onderscheiden bij de volwassen exemplaren. De meeste grote exemplaren hebben een lange rechte schedelkam die schuin naar achteren wijst en soms zelfs in een lichte kromme wat naar beneden buigt. Sommige specimina echter hebben een kam die boven de hersenpan sterk omhoog buigt en zelfs naar voren kromt. Daarna buigt die dan plots naar achter, vormt een holle bovenkant, buigt even weer naar boven in een uitstekend puntje en gaat dan vrij verticaal naar beneden zodat het geheel een kortere opstaande structuur vormt. Een ander onderscheid is dat de bek sterker naar boven buigt; dit is wel gezien als een aanwijzing dat zich daar een voorste hoornkam zou kunnen bevinden, wat ook de hoge en alleen bewaard gebleven achterkant van de snuit van een beschadigd exemplaar, UALVP 24238, suggereert. Verder lijkt het achterhoofdsgat zich meer naar onder te bevinden zodat de kop meer naar beneden gericht gehouden werd en lopen de praemaxillae meer door naar achteren. De bekendste schedel van dit type werd door Harksen P. sternbergi gedoopt. Verschillen in de postcrania zijn nooit gevonden. Deze morfen zouden zich volgens Bennett echter niet gelijktijdig in de populatie voordoen; dat van P. sternbergi zou alleen in de oudste aardlagen van de Niobrara gevonden zijn, terwijl het andere type pas honderdduizenden jaren later opduikt en voorkomt tussen 85,8 en 83,5 miljoen jaar geleden. Hieruit concludeerde hij dat de ene morfe de voorouder was van de andere. Het zou bij elkaar dus om een doorlopende reeks chronospecies gaan; wegens het duidelijke verschil in bouw besloot Bennett een taxonomisch soortonderscheid te handhaven en P. sternbergi dus als valide naam te handhaven naast P. longiceps, hoewel er bij zo'n directe afstammingsrelatie biologisch geen strikte grens te trekken zou zijn. Zou men een geslachtsonderscheid willen maken, zoals Kellner doet, dan is de naam Geosternbergia de geldige voor de oudere vorm.

Een directe afstammingsrelatie is moeilijk te bewijzen en wordt ook maar zelden aangenomen. Juist het beste bewijs ervoor: een doorlopende reeks overgangsvormen die een geleidelijke gradiënt zou tonen van de ene naar de andere vorm, zou het ook weer onmogelijk maken een onderscheid te handhaven en dan zou P. occidentalis als enige soort overblijven, waarin P. longiceps en P. sternbergi zouden samenvallen. Een probleem is daarbij dat vroegere onderzoekers de stratificatie niet nauwgezet bijhielden zodat niet bekend is of P. longiceps toch niet eerder voorkwam. Verder is het holotype van P. sternbergi, verreweg het beste materiaal dat ondubbelzinnig van de soort bekend is en het enige dat de kamvorm goed toont, de grootste bekende pteranodonschedel, die zou kunnen wijzen op een spanwijdte van acht meter; de oorspronkelijke beschrijvers gingen zelfs uit van negen meter. De onderkaaklengte ervan is 1217 millimeter en de kam tachtig centimeter hoog. Dat maakt het echter mogelijk dat deze morfe geen aparte soort is maar de vorm vertegenwoordigt waartoe uitzonderlijk grote, misschien mannelijke, individuen uitgroeiden. David Unwin heeft erop gewezen dat zelfs onder de grootste exemplaren van Bennetts populatieonderzoek zich nog onvolwassen individuen bevonden. Volgens Bennett zijn individuen uit de diepere en dus oudere aardlagen daarbij gemiddeld iets kleiner, wat het holotype nog uitzonderlijker maakt.
In Kelners interpretatie van 2010 zijn er juist meerdere parallelle afstammingslijnen mogelijk. Geosternbergia sternbergi is wellicht de voorouder van Geosternbergia maiseyi; Dawndraco, slechts bekend van het afwijkende specimen UALVP 24238, zou de voorouder geweest kunnen zijn van Pteranodon longiceps. Wegens de vele onzekerheden over welke variatie nu individueel, leeftijdsbepaald of seksebepaald is, achtte Kelner het onmogelijk om aan Geosternbergia sternbergi, Geosternbergia maiseyi of Dawndraco meer fossielen toe te wijzen dan hun holotypen.
Unieke kenmerken van Pteranodon longiceps
[bewerken | brontekst bewerken]Volgens Kellner is alleen P. longiceps een echte soort van Pteranodon. In dat geval zouden de unieke afgeleide kenmerken ofwel autapomorfieën van die soort en dat geslacht moeten samenvallen. In 2010 gaf hij een reeks van zulke eigenschappen waarin Pteranodon zich van Geosternbergia en Dawndraco onderscheidt. De voorhoofdskam is groot, langgerekt en naar achteren gericht. De voorrand van die kam wordt gevormd door een uitloper van de praemaxilla, het voorste been van de bovenkaak, die tot boven de oogkas doorloopt. Die voorrand maakt een hoek van vijfentwintig à dertig graden met de kaaklijn, Vóór de oogkas loopt de bovenrand van de schedel vrijwel horizontaal. De symfyse van de onderkaken, de zone waarin ze vooraan samengegroeid zijn, beloopt het voorste twee derde deel van de mandibula, de onderkaken bij elkaar. De processus nasalis heeft de vorm van een bult. De buitenste bovenkant van het opperarmbeen heeft een opening voor de bloedvaten.
Kellner wijkt in zijn beschrijving vooral van eerdere onderzoekers af in het belang dat hij hecht aan de lengte van de praemaxilla waarvan hij aanneemt dat die ver naar achteren doorloopt; Bennett nam nog aan dat de voorrand van de kam zelf alleen door het voorhoofdsbeen gevormd wordt. Door de hechte vergroeiingen is het zeer lastig precies de grens te bepalen.
Fylogenie
[bewerken | brontekst bewerken]Marsh plaatste Pteranodon in 1876 in een eigen familie Pteranodontidae en die weer in een eigen onderorde Pteranodontia. Deze indeling werd niet algemeen aanvaard. Williston gebruikte in 1893 een Ornithostomatidae en in 1897 een Ornithostomatinae als onderfamilie van de Ornithocheiridae. Miller plaatste de soort in 1971 zelfs direct in de Ornithocheiridae. Tegenwoordig wordt de soort weer algemeen in de Pteranodontidae geclassificeerd maar welke andere vormen tot die groep behoren is dan onzeker; soms worden ook Nyctosaurus of Ornithostoma als pteranodontiden gezien; soms is Pteranodon het enige lid, wat het hele begrip overbodig zou maken. Miller dacht dat Nyctosaurus de directe voorouder van Pteranodon was, een hypothese die verlaten is; zoals meestal kennen we ook van Pteranodon de vooroudersoort niet.
Het vraagstuk van de precieze afstamming van de pterosauriërs is de afgelopen vijftien jaar meer exact bestudeerd door computerprogramma's de meest waarschijnlijke stamboom te laten uitrekenen. Volgens zo'n studie uit 1994 door Bennett bestaat er een klade, aftakking of afstammingsgroep, Pteranodontidae, die dus wel dezelfde naam draagt als de familie maar daarvan begripsmatig onderscheiden moet worden. De klade Pteranodontidae zou behalve Pteranodon zelf, ook Ornithodesmus, Santanadactylus en Anhanguera omvatten. Deze Pteranodontidae zouden uiteindelijk deel uitmaken van de ruimere Eupterodactyloidea. Bennets stamboom is echter achterhaald door veel uitgebreidere analyses.
Tegenwoordig zijn twee veronderstelde stambomen het invloedrijkst: die gevonden door Unwin en een alternatieve gevonden door Alexander Kellner. Beide werden gepubliceerd in 2003. Volgens Unwin is er een klade Pteranodontia die zowel Pteranodon als Nyctosaurus omvat. Deze zou nauw verwant zijn aan de Ornithocheiridae binnen de Euornithocheira. Volgens de kladistische analyse van Kellner echter is Nyctosaurus niet speciaal aan Pteranodon verwant. Pteranodon zou in Kellners fylogenie binnen de Pteranodontoidea basaal, dus onder in de stamboom, geplaatst zijn in een klade waarin hij het zustertaxon is van een meer beperkte groep waarvan weer Istiodactylus (de nieuwe naam voor Ornithodesmus), en een klade bestaande uit Ornithocheirus en de Anhangueridae deel uitmaken. Zeer verwarrend is hierbij dat de auteurs verschillende namen geven aan inhoudelijk dezelfde groepen maar juist dezelfde namen aan verschillend gedefinieerde kladen.
Volgens Unwin zag de evolutionaire stamboom, ofwel het kladogram er dus zo uit:
| Euornithocheira |
| ||||||||||||
Het cladogram van Kellner echter is het volgende:
| Pteranodontoidea |
| ||||||||||||
Fundamenteler dan hun geschil over welke de nauwste verwanten van Pteranodon zijn, is de tegenstelling tussen Unwin en Kellner aangaande de vraag tot welke ruimere groep binnen de Pterodactyloidea Pteranodon behoorde. Unwin stelt dat dit de Ornithocheiroidea zijn, op te vatten als de meest basale groep in de pterodactyloïden. In dat geval is de tandeloze snavel van Pteranodon een geval van convergente evolutie, een niet op afstamming gebaseerde gelijkenis, met de latere Azhdarchidae en is zijn zwevende visetende levensstijl misschien oorspronkelijk voor de hele groep van de Pterodactyloidea. Kellner echter vond in zijn analyse dat Pteranodon tot dezelfde Dsungaripteroidea behoorde als de azhdarchiden, zodat tandeloosheid een oorspronkelijk dsungaripteroïde kenmerk geweest zou zijn en aangenomen moet worden dat verschillende deelgroepen van de Dsungaripteroidea opnieuw tanden ontwikkelden.
Kellners splitsing van Pteranodon in 2010 verandert weinig aan de meer omvattende fylogenie; alleen moet waar hij eerst Pteranodon gebruikte, nu "Pteranodontidae" gelezen worden, welke klade hij opnieuw definieerde. De interne afstammingsrelaties binnen de Pteranodontidae gaf Kellner in 2010 nog niet aan.
Levenswijze
[bewerken | brontekst bewerken]Ter zee: de jacht
[bewerken | brontekst bewerken]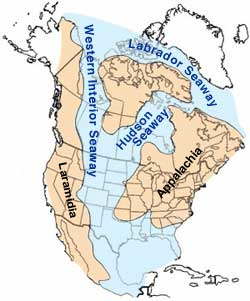
Pteranodon wordt algemeen gezien als een gespecialiseerde viseter — en dus roofdier — en zwever. Hij zou dus dezelfde rol vervuld hebben als de huidige albatros, waaraan ook zijn vliegvermogen lijkt te zijn aangepast. Als bewijs dat hij vissen at, wordt vaak verwezen naar een fossiel, AMNH 5098, dat zelfs braakballen met vissenbotten nog in de bek had, maar in dit soort mariene afzettingen zijn resten van vissen algemeen en het verband kan dus puur toevallig zijn. De fossielen zijn gevonden in de mariene lagen die afgezet zijn in de enorme binnenzee, de Western Interior Seaway, die toen van het gebied van de huidige Golf van Mexico tot in Canada liep. De vindplaatsen liggen zo'n honderdvijftig kilometer van de toenmalige kustlijn verwijderd, dus Pteranodon kon aanzienlijke afstanden over open water afleggen en was geen obligate 'kustpterosauriër'. In de formatie vertegenwoordigt hij zevenennegentig procent van de gevonden pterosauriërfossielen; de overige drie procent neemt Nyctosaurus voor zijn rekening. Deze kleinere soort kwam dus in ieder geval gelijktijdig in hetzelfde gebied voor. Dat beiden aparte niches opvulden, is waarschijnlijk maar het is niet helemaal duidelijk welke. Misschien kon Nyctosaurus niet zo heel diep de zee op.
Resten van Pteranodon zijn niet noordelijker aangetroffen dan Wyoming en South Dakota, hoewel de mariene kalkafzettingen tot in Canada doorlopen en de binnenzee wellicht tot in de poolstreek reikte. Bennett heeft deze beperking ooit gezien als een aanwijzing dat de stofwisseling van de soort niet al te hoog lag en hij zich daarom niet op hogere breedten kon handhaven. Het kan echter ook een effect zijn van lokale conserveringsomstandigheden; pterosauriërfossielen zijn normaal erg zeldzaam.
Hoe Pteranodon precies vissen of inktvissen ving, is niet zeker. Meestal wordt aangenomen dat hij laag over het water zweefde en dan met zijn korte verticaal flexibele nek en lange bek zijn prooi uit het water plukte. Lange tijd met de snavel door het wateroppervlak scheren, een methode die sommige huidige vogels, de schaarbekken, toepassen, zou voor zo'n grote soort te veel energie gekost hebben, tenminste als hij op eigen kracht de golven had moeten doorklieven; misschien dat winddruk een deel van de energie had kunnen leveren. Een alternatieve hypothese van Niall Mateer uit 1975 dat het dier op deze wijze plankton ving omdat het zonder tanden geen vissen had kunnen vasthouden, wordt daarmee extra onwaarschijnlijk. Of Pteranodon ook kon zwemmen of zelfs duiken is onbekend. Zeker was het uitgesloten dat hij zich van grotere hoogte in het water stortte, want dat zou al zijn botten gebroken hebben, maar Bennett heeft gesuggereerd dat van lagere hoogte toch een beperkte duik vanuit de vlucht mogelijk was. Vroeger werd vanwege de keelzak vaak gesteld dat Pteranodon als een pelikaan zwemmend vissen uit het water schepte, maar de keelzak, indien al aanwezig, lag daarvoor te ver naar achteren; de onderkaak is vooraan te smal om deze vangstmethode mogelijk te maken. Duiken en zwemmen zou vergemakkelijkt worden als er vliezen tussen de tenen zaten; dat die langer waren dan bij de ornithocheiriden is wel gezien als een aanwijzing voor een grotere rol van die bewegingswijzen. Een studie van Amy Smith uit 2007 toonde aan dat de klauwen van Pteranodon wel lijken op die van aan het wateroppervlak zwemmende vogels zoals meeuwen maar niet op die van echte duikvogels zoals alken. Vermoedelijk kon Pteranodon wel van het wateroppervlak opstijgen, op zelfde wijze als van de grond zich met de vleugels afzettend, hoewel in dit geval misschien in een paar sprongen om zich eerst gedeeltelijk aan de zuiging te onttrekken.
De zeeën die Pteranodon beviste, waren ook de habitat van andere amniote gewervelden: zeeschildpadden als Toxochelys en de reusachtige Archelon, plesiosauriërs als Elasmosaurus en de niet-vliegende duikvogels Baptornis en Hesperornis. De grootste zeereptielen waren de mosasauriërs Clidastes en Tylosaurus. Dit soort roofdieren joeg getuige braakballen met pterosauriërresten ook weer op Pteranodon, een aanwijzing dat die toch op het wateroppervlak neerstreek, hoewel ook wel is aangenomen dat plesiosauriërs met hun lange nekken de vliegende reptielen uit de lucht plukten. In de zee leefden ook grote roofvissen zoals haaien en Saurodontidae; in 2018 werd een fossiel beschreven dat erop wees dat zulke dieren het karkas van een pteranodon hadden aangevreten.
Ter land: voortplanting
[bewerken | brontekst bewerken]Misschien dat Pteranodon ook op het land naar voedsel zocht en vliegend binnenwateren beviste of staande aan de waterkant de prooi verschalkte. Daarvoor zijn echter geen directe bewijzen. In ieder geval moest het dier vaste grond opzoeken om zich voort te planten. We weten dat pterosauriërs in het algemeen eierleggend waren en er is geen reden aan te nemen dat Pteranodon op dit punt een uitzondering vormde. De leerachtige eieren hadden een dunne schaal en moesten met planten of grond bedekt worden om niet uit te drogen. Datzelfde plantenmateriaal zou door broeiing ook voldoende warmte hebben kunnen opgewekt om een fatale afkoeling van de embryo's te voorkomen. Daarvoor was het dus niet nodig dat een van de ouders regelmatig op het nest zat. Pasgeboren jongen van Pteranodon zijn nog niet beschreven, maar van andere pterosauriërsoorten weten we dat hun jongen al meteen bij het uitkomen voldoende grote vleugels hadden om te kunnen uitvliegen. Echte broedzorg was dus niet nodig. Als Bennett gelijk heeft met zijn hypothese over de seksuele dimorfie bemoeiden in ieder geval de mannetjes zich daar niet mee: die hadden het dan te druk met het bevruchten van zo veel mogelijk wijfjes die ze niet individueel in de opvoeding konden ondersteunen. Wel was het voordelig broedkolonies te vormen. Dat verminderde de kans dat roofdieren het nest leegvraten; de verdediging daartegen kan een van de belangrijkste redenen zijn geweest dat het wijfje na de leg in de buurt bleef. Eilanden ver uit de kust en zonder grote predatoren zullen de favoriete vestigingsplaatsen voor zo'n kolonie zijn geweest. In 2023 stelde David Hone dat de grote omvang van Pteranodon niet bereikt had kunnen worden zonder vrij lange broedzorg.
De jongste individuen waren kennelijk niet in staat de diepere zeeën te bereiken waar wij later hun fossielen konden vinden. Volgens Bennett duidt dit toch op broedzorg en waren de jongen afhankelijk van voedsel dat hun moeders brachten. Hoe snel ze daardoor groeiden is onbekend. Vermoedelijk konden ze niet in één seizoen een volwassen grootte bereiken en deden individuen die een maximale spanwijdte bereikten daar vele jaren over. Bennetts populatieonderzoek toonde aan dat onvolwassen dieren, te herkennen aan een onvolkomen vergroeiing van de beennaden en een ongelaagde botopbouw, in alle grootteklassen voorkwamen. Hij zag dat als een aanwijzing dat ze niet groter meer zouden worden en Pteranodon dus net als de vogels een groeistop kende: bij een bepaalde leeftijd of grootte zou een dier met groeien ophouden, na eerst juist erg snel gegroeid te zijn. Unwin daarentegen stelt dat als er zo'n groeistop was, de volwassen afmetingen veel minder variatie zouden tonen. Subadulte dieren zouden juist hun groei weliswaar vertraagd maar nog niet beëindigd hebben en er moeten dan volwassen reuzen zijn geweest die wij door hun relatieve zeldzaamheid nog niet hebben gevonden. In dat geval leek de groei van Pteranodon meer op die van de huidige koudbloedige reptielen.
In 2017 beschreef Bennett het tot dan toe kleinste gevonden exemplaar van Pteranodon, een vleugel die wees op een spanwijdte van 176 centimeter. Hij stelde dat het een jong moest zijn geweest. Het verschilde in proporties niet van volwassen exemplaren. Omdat het in een laag aangetroffen werd die indertijd ver uit de kust lag, zou het daar op eigen kracht naar toe moeten zijn gevlogen. Dit bewees weer dat ze toch niet van broedzorg afhankelijk waren. Het normaliter ontbreken van zulke jonge dieren in deze lagen werd nu door hem gezien als een aanwijzing dat ze een andere ecologische niche innamen dan volwassenen en wellicht dichter bij de kust hun voedsel zochten. De verschillende leeftijdsklassen fungeerden dus als ecospecies. Hier leidde Bennett uit af dat voor pterosauriërs in het algemeen de taxonomische diversiteit gering zou zijn geweest en dat er, mede gezien hun aanzienlijke lichaamsomvang, dus gelijktijdig weinig soorten leefden, net als bij huidige krokodillen.

Van warmbloedige soorten hebben de individuen een relatief korte levensduur, hoogstens enkele tientallen jaren. Dat heeft tot gevolg dat ze snel evolueren. Ook de soort zelf heeft dan een korte levensduur van gemiddeld een paar miljoen jaar voor vormen die stabiel genoeg zijn om door ons als type herkend te worden: hij splitst zich in verschillende soorten of wordt verdrongen door andere zich snel ontwikkelende soorten die met hem om dezelfde niche concurreren. Pteranodon was in dit verband een succesvol geslacht. Als we P. longiceps en P. sternbergi als een enkele chronospecies zien, hield die het zeker zo'n vijf miljoen jaar uit, van 88 tot 83 miljoen jaar geleden. Dan houdt het verslag dat de fossielen ons doen op. Net zoals we niet weten van welke vooroudersoort Pteranodon zich afsplitste, is onbekend of hij daarna zelf in aparte soorten gesplitst is of zonder nageslacht achter te laten is uitgestorven. Het is zeer onwaarschijnlijk dat de soort het tot het einde van het Krijt uithield, toen 66 miljoen jaar geleden alle grote soorten door een catastrofe, tijdens de K-T-overgang, werden uitgeroeid, wellicht door een meteorietinslag.
Volkscultuur
[bewerken | brontekst bewerken]Pteranodon gold lange tijd als grootste vliegende dier uit de wereldgeschiedenis en verwierf zich hiermee een vaste plaats in de populair-wetenschappelijke (kinder)boeken. Zelfs toen in 1975 de ontdekking van Quetzalcoatlus bekendgemaakt werd, drong het bestaan daarvan maar langzaam tot het grote publiek door. Dat Pteranodon een Amerikaanse soort is, droeg nog aan de verbreiding van zijn naam binnen de massacultuur bij. Pteranodon werd hierdoor verreweg de bekendste pterosauriër, die een standaardelement geworden is in verbeeldingen van 'De Oertijd'. Een afbeelding van de 'Tijd der Reptielen' is niet compleet zonder een paar pteranodontia in de lucht. In talloze stripverhalen, cartoons en films duikt het dier op en het is zo tot het stereotype geworden van de pterosauriër waarvan de naam veel beter gekend wordt dan die van de Pterosauria als geheel. Hierdoor is echter wel een heel fout beeld geschapen van zowel het dier zelf als van zijn verwanten. Pteranodon was niet zo typisch voor een pterosauriër. De meeste soorten waren veel kleiner en hadden geen lange kam op het achterhoofd maar wel tanden. Dat de tandeloosheid uitzonderlijk is, werd onbewust gereflecteerd door veel speelgoedmodellen van Pteranodon die door de makers alsnog van een vervaarlijk gebit werden voorzien. Toen in de jaren negentig een pterosauriër aan het licht kwam die echt een lange kam met tanden combineerde, werd deze door de ontdekkers dan ook ironisch Ludodactylus gedoopt, 'speelgoedpterodactyl'. Ook wordt er een foute suggestie gegeven van de verspreiding in ruimte en tijd. Pteranodon leefde weliswaar miljoenen jaren lang maar die vormden maar een kort deel van het hele Mesozoïcum. Ook was zijn leefgebied wellicht erg beperkt. Het overgrote deel van de dinosauriërs heeft nooit een pteranodon kunnen zien als ze omhoog keken.
Literatuur
- Newberry, J. S., 1871, "Announcement of discovery, by Prof. O. C. Marsh, of a species of Pterodactyl", Proc. Lyc. nat. Hist. N. Y. 1: 241
- Marsh, O. C., 1876, "Notice of a new sub-order of Pterosauria", American Journal of Science, Series 3, 11: 507-509
- Delafontaine, M., 1877, "Remarks on pteranodonts", Arch. Sci. Phys. Nat. 49: 127
- Marsh, O. C., 1884, "Principal characters of American Cretaceous pterodactyls. Part 1. The skull of Pteranodon", American Journal of Science, Series 3, 27: 423-426
- Dollo, L., 1888, "Sur la signification de la créte occipitale de Pteranodon", Proc.-verb. Soc. belg. Géol. Paléont. Hydrol. 2: 79–80
- Williston, S. W., 1891, "The skull and hind extremity of Pteranodon", The American Naturalist, 25: 1124-1126
- Williston, S. W., 1895, "Note on the mandible of Ornithostoma", Kansas University Quarterly, 4:61
- Williston, S. W., 1896, "On the skull of Ornithostoma", Kansas University Quarterly, 4: 195-197
- Williston, S. W., 1897, "Restoration of Ornithostoma (Pteranodon)", Kansas University Quarterly, 6: 35-51
- Eaton, G. F., 1903, "The characters of Pteranodon", American Journal of Science, 16: 82-86
- Eaton, G. F., 1904, "Characters of Pteranodon", American Journal of Science, 17: 318-320
- Eaton, G. F., 1908, "The skull of Pteranodon", Science 27: 254–255
- Eaton, G. F., 1910, "Osteology of Pteranodon", Memoirs of the Connecticut Academy of Arts and Sciences, 2: 1-38
- Hankin E.H. & Watson D.S.M., 1914, "On the Flight of Pterodactyls", The Aeronautical Journal, October 1914, pp. 324–335
- Gilmore, C. W., 1935, "A new occurrence of the flying reptile, Pteranodon", Science, 82: 371
- Kripp, D. von, 1943, "Ein Lebensbild von Pteranodon ingens auf flugtechnischer Grundlage", Nova Acta Leopoldina, N.F., 12(83): 16-32
- Traatz, E. P., 1955, "Eine lebensgrosse Nachbildung des Pteranodon", Der Präparator, 2: 113-117
- Enlow, D. H. and Brown, S. O., 1958, "A comparative histological study of fossil and recent bone tissues. Part III", Texas Journal of Science, 10: 187-230
- Sternberg, G. F. and Walker, M. V., 1958, "Observation of articulated limb bones of a recently discovered Pteranodon in the Niobrara Cretaceous of Kansas", Transactions of the Kansas Academy of Science, 61: 81-85
- Harksen, J. C., 1966, "Pteranodon sternbergi, a new pterodactyl from the Niobrara Cretaceous of Kansas", Proceedings, South Dakota Academy of Science, 45: 74-77
- Bramwell, C. D. and Whitfield, G. R., 1970, "Flying speed of the largest aerial vertebrate", Nature, 225: 660
- Bramwell, C. D., 1971, "Aerodynamics of Pteranodon", Biological Journal of the Linnean Society, 3: 313-328
- Heptonstall, W. B., 1971, "An analysis of the flight of the Cretaceous pterodactyl Pteranodon ingens (Marsh)", Scottish Journal of Geology, 7: 61-78
- Miller, H. W., 1972, "The taxonomy of the Pteranodon species from Kansas", Transactions of the Kansas Academy of Science, 74: 1-19
- Miller, H. W., 1972, "A skull of Pteranodon (Longicepia) longiceps Marsh associated with wing and body bones", Transactions of the Kansas Academy of Science, 74: 20-23
- Miller, H. W., 1972, "Longicepia Miller (1972) preoccupied by Pteranodon Marsh", Transactions of the Kansas Academy of Science, 75: 100
- Ikuwo, O., Hasegawa, Y. & Otsuka, H., 1972, [in het Japans] "Preliminary report on the Cretaceous reptile fossils from Hokkaido", Mem. nat. Sci. Mus. 5: 213–222
- Pinna, G., 1973, "La riconstruzione di uno pterodonte (Pteranodon ingens Marsh) esposta nel Museo Civico di Storia Naturale di Milano", Natura, 64: 35-39
- Bramwell, C. D. and Whitfield, G. R., 1974, "Biomechanics of Pteranodon", Philosophical Transactions of the Royal Society of London, B, 267: 503-581
- Mateer, N. J., 1975, "A study of Pteranodon", Bulletin of the Geological Institutions of the University of Uppsala, N.S., 6: 23-33
- Stein, R. S., 1975, "Dynamic analysis of Pteranodon ingens: a reptilian adaptation to flight", Journal of Paleontology, 49: 534-548
- Stein, R. S., 1976, "Aerodynamics of the long pterosaur wing", Science, 191: 898-899
- Whitfield, G. R., 1979, "Engineering and aerodynamics of a flying dinosaur: Pteranodon", Sci. Mech. pp. 56–58 & 118
- Sneyd, A. D., Bundock, M. S., and Reid. D., 1982, "Possible effects of wing flexibility on the aerodynamics of Pteranodon", The American Naturalist, 120: 455-477
- Brower, J. C., 1983, "The aerodynamics of Pteranodon and Nyctosaurus, two large pterosaurs from the Upper Cretaceous of Kansas", Journal of Vertebrate Paleontology, 3: 84-124
- Schoch, R. M., 1984, "Notes on the type specimens of Pteranodon and Nyctosaurus (Pterosauria, Pteranodontidae) in the Yale Peabody Museum collections", Postilla, 194: 1-23
- Winkworth, S., 1985, "Pteranodon flies again", New Scientist, 3 Jan. 1985: 32-33
- Winkworth, S., 1985, "Pteranodon", Flug und Modelltechnik 359: 990–993
- Bennett, S. C., 1987, "New evidence on the tail of the pterosaur Pteranodon (Archosauria: Pterosauria)", pp. 18–23 in: Currie, P. J. and Koster, E. H., eds. Fourth symposium on Mesozoic terrestrial ecosystems, Short papers. Occasional Papers of the Tyrrell Museum of Paleontology, 3
- Bennett, S. C., 1987, "Sexual dimorphism in the pterosaur Pteranodon", Journal of Vertebrate Paleontology, 7(Supplement to #3): 11A
- Bennett, S. C., 1988, "Pteranodon subadults, and the ontogeny of pterosaurs", Journal of Vertebrate Paleontology, 8(Supplement to #3):9A
- Bennett, S. C., 1991, Morphology of the Late Cretaceous pterosaur Pteranodon and systematics of the Pterodactyloidea
- Bennett, S. C., 1992, "Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests", Journal of Vertebrate Paleontology, 12: 422-434
- Bennett, S. C., 1993, "The ontogeny of Pteranodon and other pterosaurs", Paleobiology, 19: 92-106
- Bennett, S. C., 1994, "Taxonomy and systematics of the Late Cretaceous pterosaur Pteranodon (Pterosauria, Pterodactyloidea)", Occasional Papers of the Museum of Natural History, University of Kansas, Lawrence, 169: 1-70
- Bennett, S. C., 1994, "The pterosaurs of the Niobrara Formation", The Earth Scientist, 11(1): 22-25
- Bennett, S. C., 2001, "The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon", Palaeontographica, A 260: 1-153
- Smith, A.C., 2007, Pteranodont claw morphology and its implications for aquatic locomotion
- Hargrave, J. E., 2007, "Pteranodon (Reptilia: Pterosauria): Stratigraphic distribution and taphonomy in the lower Pierre Shale Group (Campanian) western South Dakota and eastern Wyoming", pp. 215–225 in: Martin, J. E. and Parris, D. C. eds. The Geology and Paleontology of the Late Cretaceous marine deposits of the Dakotas, The Geological Society of America, Special Paper 427
- Bennett, S.C., 2009, "Morphological evolution of the wing of pterosaurs: myology and function", Zitteliana B 28: 127 - 141
- Kellner, A.W.A., 2010, "Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species", Anais da Academia Brasileira de Ciências 82(4): 1063-1084
- S. Christopher Bennett and Paul Penkalski, 2017, "Waves of bone deposition on the rostrum of the pterosaur Pteranodon", Geological Society SP455. New Perspectives on Pterosaur Palaeobiology doi:10.1144/SP455.2
- S. Christopher Bennett, 2017, "New smallest specimen of the pterosaur Pteranodon and ontogenetic niches in pterosaurs", Journal of Paleontology DOI: https://doi.org/10.1017/jpa.2017.84Published
- Dana J. Ehret & T. Lynn Harrell, Jr., 2018, "Feeding traces on a Pteranodon (Reptilia: Pterosauria) bone from the Late Cretaceous (Campanian) Mooreville Chalk in Alabama, USA". Palaios 33(9): 414–418
- Yang, Z.; Jiang, B.; Benton, M. J.; Xu, X.; McNamara, M. E. & Hone, D. W. E. 2023. "Allometric wing growth links parental care to pterosaur giantism". Proceedings of the Royal Society B: Biological Sciences. 290(2003): 20231102
Noten
- ↑ Pteranodon. www.prehistoric-wildlife.com. Gearchiveerd op 21-2-2023. Geraadpleegd op 30-12-2022.
- ↑ Fossilworks: Pteranodon. www.fossilworks.org. Geraadpleegd op 30-12-2022.
- ↑ Dixon, Dougal (2001). Dinosauriërs In De Lucht. Corona Gidsen, p. 18. ISBN 9054955945.