Kekapis
| Kekapis | |
|---|---|
| |
| Pengelasan saintifik | |
| Alam: | |
| Filum: | |
| Kelas: | |
| Order: | |
| Keluarga: | Pectinidae
|
| genus | |
|
Rujuk teks | |
| Sinonim | |
|
Pectenidae | |
Kekapis atau kapis ialah moluska bivalvia (dwicangkerang) marin yang tergolong dalam keluarga taksonomik Pectinidae.
Kekapis merupakan sebuah keluarga bivalvia kosmopolitan yang terdapat di kesemua lautan dunia tetapi tidak sekali pun di air tawar. Ia merupakan antara sekumpulan kecil bivalvia yang "hidup bebas", yang mana kebanyakan spesiesnya mampu berenang dengan deras dalam jarak pendek, malah bermigrasi cukup jauh merentasi dasar lautan. Sebilangan kecil spesies kapis hidup terlekap pada substrat (tapak) batuan ketika dewasa, dan ada lagi yang melekat pada objek pegun atau berakar rumput laut pada suatu ketika dalam hidup dengan menggunakan filamen rembesannya yang dipanggil benang bisus. Majoriti spesies kapis pula hidup secara berbaring di atas tapak pasir, dan apabila terasa kehadiran pemangsa seperti tapak sulaiman, ia boleh cuba melarikan diri dengan berenang secara cepat dan lintang-pukang melalui pendorongan jet yang terhasil dengan mengepak-ngepakkan cangkerangnya. Kapis mempunyai sistem saraf yang cukup berkembang, malah ia berbeza dengan bivalvia lain di mana kesemua spesies kapis mempunyai deretan mata-mata ringkas yang membarisi bibir mantelnya (selaput di bawah cangkerang).
Kebanyakan spesies kapis amat dihargai sebagai sumber makanan manusia, malah ada juga yang dijadikan ternakan akuakultur. Perkataan "kapis" (bahasa Inggeris: scallop) juga merujuk kepada dagingnya, iaitu otot aduktor, yang dijual sebagai makanan laut. Cangkerang kapis yang berwarna-warni, bersimetri dan berbentuk kipas, lebih-lebih lagi dengan corak jalur-jaluran yang memancar, dihargai oleh pengumpul cangkerang dan digunakan sebagai motif seni halus, seni bina dan reka bentuk semenjak zaman-berzaman.

Biologi
[sunting | sunting sumber]Taburan dan habitat
[sunting | sunting sumber]Kekapis mendiami semua lautan di dunia, dengan kawasan Indo-Pasifik mencatatkan jumlah spesies penghuni yang tertinggi. Kebanyakan spesies tinggal di perairan yang agak cetek dari paras air surut hingga 100 m, sedangkan yang lain gemar air dalam. Biarpun sesetengah spesies hidup di persekitaran yang amat sempit sahaja, tetapi kebanyakannya bersifat oportunis dan boleh hidup dalam pelbagai ragam keadaan. Kekapis boleh ditemui hidup-hidup di celah-celah, atas atau bawah batu-batuan, karang, kerikil, rumput laut, kelpa, pasir, atau lumpur. Kebanyakan kapis memulakan hayat sebagai anak-anakan berlekap bisus, iaitu kebolehan yang dipelihara oleh sesetengah kapis sepanjang hidup walhal yang lain tumbuh menjadi dewasa yang hidup bebas.[1]
Anatomi dan fisiologi
[sunting | sunting sumber]Sedikit sahaja variasi yang berlaku dalam susunan organ dan sistem dalaman dalam kalangan keluarga kapis, maka keterangan yang berikutnya boleh diperlakukan pada sebarang spesies kapis.[2]
Orientasi
[sunting | sunting sumber]
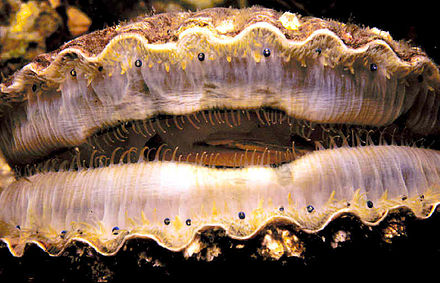
Cangkerang kapis terdiri daripada dua belah atau "injap" (valves), iaitu satu kiri dan satu kanan, yang terbahagi dengan satah simetri.[3] Kebanyakan spesies kapis berbaring pada injap kanan, dan oleh itu injap kanan selalunya lebih jeluk dan bundar daripada injap kiri (atas) yang sebenarnya berbentuk cekung pada kebanyakan spesies. Engsel yang menghubungkan kedua-dua injap berorientasi ke arah atas, maka dari segi morfologi, setengah kapis bersamaan dengan bahagian anterior atau hadapan, dan setengah lagi ialah posterior atau belakang, manakala engselnya merupakan dorsal atau belakang atas, dan bahagian bawah pula merupakan ventral atau perutnya.[4] Akan tetapi, oleh sebab kebanyakan cangkerang kapis bersifat simetri bilateral atau "injap sekata", apatah lagi bersimetri hadapan ke belakang atau "sisi sekata", maka penentuan "muka" kapis memerlukan maklumat terperinci tentang injap-injapnya.
Injap
[sunting | sunting sumber]
Cangkerang kapis yang misali terdiri daripada dua injap yang berbentuk seiras dengan garis engsel yang lurus sepanjang atasnya, tiada gigi, dan menghasilkan sepasang sayap pipih atau "telinga" (juga dipanggil "aurikel" tetapi ini juga istilah yang digunakan untuk dua kebuk di dalam jantungnya) di mana-mana belah dari titik tengahnya, satu ciri yang unik dan kelihatan pada kesemua kapis dewasa.[5] Mungkin kedua-dua telinga sama saiz dan bentuk, dan mungkin juga telinga anterior lebih besar (telinga posterior tidak pernah lebih besar daripada yang anterior, dan inilah ciri penting untuk menentukan yang mana injap kiri dan kanan).
Seperti pada hampir kesemua jenis bivalvia, terdapat garis-garis dan/atau gegelang pertumbuhan yang berpunca di tengah engsel, pada titik yang dipanggil "paruh" (beak) yang dikelilingi oleh kawasan yang umumnya tertimbul dipanggil umbo atau "bonggol". Gelang pertumbuhan ini membesar ke bawah sehingga mencapai tepi cangkerang ventral yang melengkung. Kebanyakan kapis mempunyai cangkerang yang bergaris arus (streamline) untuk memudahkan pergerakan ketika berenang pada suatu ketika dalam kitaran hidupnya, sambil juga memperoleh perlindungan daripada pemangsa. Kapis yang injapnya berabung mempunyai kelebihan dari segi binaan yang diperoleh daripada rabung-rabung tersebut iaitu "tetulang", namun tetulang itu agak menambah berat dan jisim.
Satu ciri yang unik terdapat pada keluarga kapis ialah kehadiran struktur seakan sikat dipanggil ctenolium yang terletak di hujung anterior injap kanan bersebelahan dengan takuk bisus injap tersebut, suatu ciri cangkerang yang amat ketara dan penting dari segi taksonomi. Kesemua kapis mempunyai ctenolium di suatu ketika dalam hayatnya biarpun kebanyakannya akan tersingkir ciri ini apabila menjadi dewasa yang bebas berenang, malah tiada bivalvia lain yang mempunyai ciri cangkerang seumpamanya. Bahagian ctenolium terdapat kapis moden sahaja; ia tidak terjumpa pada kedua-dua calon leluhur kepada kapis moden, iaitu Entoliidae dan Aviculopectinidae.[6]
Sistem otot
[sunting | sunting sumber]
Sama seperti tiram (keluarga Ostreidae), kapis mempunyai sebatang otot aduktor pusat; oleh itu, bahagian dalam cangkerangnya terdapat parut pusat yang menandakan tempat melekapnya otot tersebut. otot aduktor kapis lebih besar dan berkembang daripada tiram kerana kapis aktif berenang; malah sesetengah spesies kapis diketahui bergerak secara berkumpulan dari satu tempat ke satu tempat. Cangkerang kapis selalunya berbentuk sekata dan sering dijadikan contoh bentuk cangkerang laut yang terulung.[4]
Sistem pencernaan
[sunting | sunting sumber]Kapis merupakan pemakan menapis yang makan plankton. Tidak seperti kebanyakan bivalvia lain, ia kekurangan bahagian sifon. Air bergerak melalui struktur penapis di mana zarah-zarah makanan terperangkap dalam mukus. Kemudian, silium-silium pada struktur tersebut menggerakkan makanan ke arah mulut. Kemudian, makanan dicerna dalam kelenjar pencernaan, iaitu organ yang kadang-kadang digelar sebagai "hati" secara kurang tepatnya, yang menyelaputi sebahagian esofagus dan usus dan seluruh perut. Bahan kumuhan disalurkan melalui usus (seperti moluska lain, bahagian hujung ususnya masuk dan keluar dari jantung) dan keluar melalui dubur.[4]:p.20
Sistem saraf
[sunting | sunting sumber]
Seperti semua bivalvia, kapis tiada otak. Sistem sarafnya dikawal oleh tiga pasang ganglion yang terletak di pelbagai titik sepanjang anatominya, iaitu ganglion serebrum atau cerebropleural, ganglion pedal, dan ganglion visera atau parietovisceral; kesemuanya berwarna kekuningan.
Ganglion visera adalah ganglion yang paling besar dan meluas, dan hadir sebagai gumpalan yang hampir terlakur hampir di tengah-tengah kapis – secara perkadaran, inilah set ganglia yang terbesar dan paling rumit dalam mana-mana bivalvia moden. Dari sinilah terpancarnya semua saraf yang menghubungkan ganglion visera dengan gegelang saraf circumpallial yang mengelilingi mantel dan berhubung dengan kesemua sesungut dan mata kapis. Gegelang saraf ini begitu berkembang maju sehingga bagi sesetengah spesies, ia secara absah boleh dianggap sebagai satu ganglion tambahan.[4]:p.46 Dari ganglion visera juga berpuncanya saraf-saraf brankia yang mengawal insang kapis.
Ganglion serebrum adalah set ganglia kedua terbesar dan terletak berjauhan dengan satu sama lain pada arah dorsal dari ganglion visera. Ia tersambung dengan ganglion visera melalui tisu penghubung serebrum-visera yang panjang, dan ia tersambung dengan satu sama lain melalui komisur (silang saraf belakang) serebrum yang menjangkau secara melengkung ke arah dorsal di keliling esofagus. Ganglion serebrum mengawal mulut kapis melalui saraf palpa (cuaran deria sentuh) dan berhubung dengan statocyst yang membolehkan kapis menderia kedudukannya dalam persekitaran sekeliling. Ia tersambung dengan ganglion pedal melalui tisu penghubung serebrum-pedal yang pendek.
Ganglion pedal tidak terlakur tetapi terletak berapat-rapatan dekat garisan tengah. Dari ganglion pedal inilah kapis menghasilkan saraf pedal yang mengawal pergerakan kakinya yang kecil dan berisi, dan juga deria rasa di dalamnya.[4]:pp. 43–47
Pembiakan
[sunting | sunting sumber]Keluarga kapis agak berlainan daripada bivalvia lain di mana sesetengah ahlinya adalah diesius (jantan dan betina berasingan), yang lain pula hermafrodit serentak (kedua-dua jantina dalam satu individu), malah ada sebilangan kecil hermafrodit protoandri (mula-mula jantan, kemudian bertukar betina). Alat kelamin yang berwarna merah menandakan betina, dan yang putih menandakan jantan. Spermatozoon (dari jantan) dan ovum (telur, dari betina) dibebaskan ke dalam air ketika musim mengawan, dan ovum yang disenyawakan tenggelam ke bawah. Setelah beberapa minggu, ovum pun menetas dan larvanya menyerupai dewasa tetapi halus dan lutsinar, hanyut ke dalam kalangan plankton sehingga bertempat semula di bawah untuk membesar, biasanya melekat dengan bebenang bisus. Sesetengah kapis seperti Argopecten irradians (kapis teluk Atlantik) singkat jangka hayatnya, dan sesetengah lagi mampu hidup menjangkau 20 tahun ke atas. Umur selalunya boleh ditentukan daripada bilangan gegelang sepusat dalam cangkerang yang dipanggil anulus.[4]
Mata
[sunting | sunting sumber]

Kapis mempunyai sebilangan besar (sehingga 200 biji) mata kecil (kira-kira 1 mm) yang terderet sepanjang tepi mantelnya. Mata-mata ini menggambarkan satu kelainan bagi kapis di kalangan moluska, menggunakan hablur-hablur guanina yang berbentuk cermin parabola cekung untuk memfokuskan dan memantulkan balik cahaya, menggantikan tempat kanta yang terdapat pada pelbagai jenis mata yang lain.[8] Di samping itu, matanya mempunyai retina dua lapis, iaitu retina luar untuk bergerak balas paling cepat dengan cahaya, dan retina dalam bertindak sedemikian dengan kegelapan yang tiba-tiba.[9] Biarpun mata ini tidak boleh mentafsir rupa dengan kejituan yang tinggi, namun kepekaan gabungan kedua-dua retina kepada cahaya yang memasuki mata dan cahaya yang terpantul balik dari cermin tersebut memberikan kejelasan kontras yang cemerlang di samping keupayaan mengesan corak cahaya dan pergerakan yang berubah-ubah.[10][11] Kapis amat bergantung pada matanya sebagai sistem pengesanan ancaman awal, yang mana ia mengimbas kawasan sekitar untuk pergerakan dan bayang-bayang yang mungkin menandakan kehadiran pemangsa. Di samping itu, sesetengah kapis mengubah corak renang atau pemakanannya mengikut keruh jernih airnya, dengan mengesan pergerakan jisim berzarah dalam kolum air.[12]
Otot aduktor
[sunting | sunting sumber]
Kapis memiliki otot-otot aduktor yang cepat (berjalur) dan perlahan (licin) dengan mempunyai struktur dan sifat mengecut yang berbeza-beza. Otot-otot ini terletak berapat-rapatan seakan terberkas tetapi terpisah dengan helaian tisu penghubung. Otot aduktor berjalur mengecut sangat cepat untuk berenang, manakala otot aduktor licin mengecut dengan perlahan dan mengatupkan cangkerang tanpa banyak menggunakan tenaga.[13]
Gerak alih
[sunting | sunting sumber]

Rata-rata kekapis hidup bebas dan aktif, tidak seperti kebanyakan bivalvia yang bergerak perlahan dan hidup terbenam. Dipercayai bahawa semua kapis lahir dengan bahan bisus yang melekapkannya pada suatu bentuk substrat seperti rumput eelgrass ketika masih kecil. Kebanyakan spesies kehilangan bisus apabila membesar, malah sedikit sahaja yang akan melekat pada substrat keras (cth. Chlamys distorta dan Hinnites multirigosus).[14]
Akan tetapi, majoriti jenis kapis hidup bebas dan boleh berenang dengan pecutan yang singkat untuk melarikan diri dari pemangsa (terutamanya tapak sulaiman) dengan membuka tutup injapnya dengan cepat. Sesungguhnya, aktiviti sedemikian dibolehkan oleh segalanya tentang bentuk cangkerangnya – termasuk simetri, ruang dalam yang sempit, permukaan yang licin dan/atau beralur, engsel yang kecil dan anjal, otot aduktor yang berkuasa, dan tepinya yang selanjar dan berlengkung sekata. Kapis selalunya memecut secara pancutan beberapa saat, kemudian mengatup rapat-rapat dan tenggelam kembali ke dasar persekitarannya.
Kapis mampu bergerak melalui kolum air sama ada secara mara/arah ventral (iaitu "berenang") dengan menyedut air ke dalam celah injapnya, kemudian melepaskan air melalui liang-liang hembusan kecil yang berdekatan dengan garis engsel; ataupun secara mengundur/arah dorsal ("melompat") dengan melepaskan air ke hadapan. Kapis yang melompat biasanya mendarat pada dasar laut antara setiap kali injapnya mengecut, manakala kapis yang berenang rata-rata kekal dalam kolum airnya ketika mengecut dan melakukan perjalanan yang lebih jauh (tetapi jarang melebihi paras 1 meter dari dasar laut dan jarang melebihi jarak 5 meter).[14] Kedua-dua pegerakan ini menelan tenaga yang amat banyak, malah kebanyakan kapis hanya mampu beralih sebanyak empat atau lima kali sebelum letih sepenuhnya dan perlu berehat selama berjam-jam. Seandainya kapis berenang mendarat di sebelah kirinya, ia mampu membalik ke kanan melalui gerakan mengepakkan cangkerang yang dipanggil refleks imbangan. Ada juga yang bergelar 'kapis menyanyi' yang dikhabarkan menghasilkan bunyi pop-pop yang perlahan ketika mengepak-ngepakkan cangkerang di dalam air (namun ia tetap menjadi persoalan).[15] Ada juga kapis yang menganjurkan kakinya dari celah injap dan menggali pasir dengan mengecutkan otot kaki.[16]
Mutiara
[sunting | sunting sumber]
Ada kalanya kapis menghasilkan mutiara, tetapi mutiara kapis kekurangan lapisan lutcahaya atau "nakre" yang memberikan nilai kepada mutiara tiram tipai, malah juga ketiadaan kilauan dan pendarrona. Malah ianya mungkin suram, kecil dan bermacam-macam warna, tetapi masih ada yang dihargai kerana paras rupanya.[17]
Kesalingan
[sunting | sunting sumber]Sesetengah kapis seperti Chlamys hastata sering membawa "penumpang" seperti span dan teritip pada cangkerangnya. Hubungan span dengan kapis disifatkan sebagai sejenis kesalingan (mutualism) atau simbiosis kerana span itu memberikan perlindungan dengan mencegah kaki tiub tapak sulaiman daripada melekap,[18][19][20] menyamarkan Chlamys hastata daripada pemangsa,[19] atau membentuk rintangan di keliling bukaan bisus untuk menghalang tapak sulaiman daripada memasukkan membran pencernaannya.[20] Selaputan span juga melindungi C. hastata daripada dihinggapi larva teritip, yakni sebagai perlindungan daripada penumpang yang meningkatkan keterdedahannya kepada pemangsa. Oleh itu, larva teritip lebih sering menempatkan diri pada cangkerang yang bebas span daripada yang diselaputi span.[18]
Sebenarnya, selaputan teritip menjejaskan daya renang C. hastata kerana ia terpaksa menelan lebih banyak tenaga dan menunjukkan perbezaan ketara dalam penggunaan tenaga anaerobik jika dibandingkan dengan yang bebas teritip. Tanpa diselaputi teritip, seekor kapis mampu berenang dengan lebih lama, jauh dan tinggi.[21]
Kitaran hidup dan pertumbuhan
[sunting | sunting sumber]Kebanyakan kapis bersifat hermafrodit (khunsa, mempunyai alat kelamin betina dan jantan sekali gus), mengubah seksnya sepanjang hayat, sedangkan yang lain wujud sebagai spesies diesius yang lahir dengan jantina tertentu. Dalam hal ini, kapis dapat ditentukan jantinanya dengan melihat warna alat kelaminnya: jantan mempunyai testis putih, betina pula mempunyai ovari merah, kedua-duanya mengandungi benih (telur). Kapis biasanya mencapai keaktifan seksual pada umur dua tahun tetapi tidak banyak menghasilkan telur sehingga berumur empat tahun. Proses pembiakan berlaku secara luaran menerusi pembenihan (spawning) di mana telur dan sperma dilepaskan ke dalam air. Pembenihan biasanya berlaku pada akhir musim panas dan awal musim luruh; juga berlakunya pembenihan musim bunga di kawasan Mid-Atlantic Bight.[22] Kapis betina sangat produktif dan mampu menghasilkan beratus-ratus juta telur setahun.[22]
Telur yang disenyawakan (berpadu dengan sperma) akan menjadi plankton, iaitu mikroorganisma yang hidup berkumpulan dan hanyut dalam air tawar mahupun masin. Larvanya kekal di kolum airnya selama empat ke tujuh minggu sebelum melesap ke dasar laut di mana ia melekap pada objek melalui bebenang bisus. Bisus itu akhirnya hilang apabila kapis mendewasa, ketika hampir semua spesies kapis beralih menjadi perenang bebas. Pertumbuhan deras berlaku dalam beberapa tahun pertama dengan peningkatan ketinggian cangkerang sebanyak 50–80 peratus serta berat dagingnya meningkat empat kali ganda, sehingga mencapai saiz boleh dagang pada umur empat hingga lima tahun.[22] Sesetengah kapis diketahui mencapai umur sebanyak 20 tahun ke atas.[23]
Taksonomi dan filogeni
[sunting | sunting sumber]Etimologi
[sunting | sunting sumber]Nama keluarga Pectinidae yang berdasarkan nama genus tipnya iaitu Pecten, datang dari bahasa Latin iaitu pecten yang bermaksud sikat, merujuk pada struktur seakan sikat pada cangkerang yang terletak di sebelah takuk bisus.[24]
Filogeni
[sunting | sunting sumber]
Sejarah fosil kapis kaya dengan spesies dan spesimennya. Rekod terawal diketahui tentang kapis sejati (yang mempunyai ctenolium) terdapat dari zaman Trias, iaitu lebih 200 juta tahun dahulu.[6] Spesies-spesies terawal terbahagi kepada dua kumpulan: satu dengan permukaan luaran yang hampir licin, iaitu Pleuronectis (von Schlotheim, 1820); dan satu lagi dengan tetulang memancar dan aurikel, iaitu Praechlamys (Allasinaz, 1972).[25] Rekod fosil juga menunjukkan bahawa kepelbagaian spesies dalam Pectinidae telah banyak berubah dari zaman ke zaman; Pectinidae merupakan keluarga bivalvia yang paling pelbagai pada zaman Mesozoik, tetapi ia nyaris lenyap sepenuhnya pada hujung zaman Batu Kapur. Ahli-ahli yang terselamat cepat berpecah menjadi pelbagai spesies pada zaman Tertier. Hampir 7,000 nama spesies dan subspesies telah diperkenalkan untuk Pectinidae fosil mahupun terkini.[26]
Kladogram yang berikut dihasilkan berdasarkan filogeni molekul menggunakan penanda gen mitokondrion (12S, 16S) dan nukleus (18S, 28S, H3) oleh Yaron Malkowsky dan Annette Klussmann-Kolb pada 2012.[27]
| Pteriomorphia |
| ||||||||||||||||||||||||||||||||||||||||||



Senarai subkeluarga dan genus
[sunting | sunting sumber]

Yang berikut adalah golongan-golongan yang teriktiraf dalam keluarga Pectinidae:
- Subkeluarga Camptonectinae Habe, 1977[28]
- Camptonectes Agassiz, 1864
- Ciclopecten Seguenza, 1877
- Delectopecten Stewart, 1920
- Hyalopecten A. E. Verrill, 1897
- Pseudohinnites Dijkstra, 1989
- Sinepecten Schein, 2006
- Subkeluarga Palliolinae Korbkov in Eberzin, 1960
- Suku Adamussiini Habe, 1977
- Adamussium Thiele, 1934
- Antarctipecten Beu & Taviani, 2013 †
- Duplipecten Marwick, 1928 †
- Lentipecten Marwick, 1928 †
- Leoclunipecten Beu & Taviani, 2013 †
- Ruthipecten Beu & Taviani, 2013 †
- Suku Eburneopectinini T. R. Waller, 2006 †
- Eburneopecten Conrad, 1865 †
- Suku Mesopeplini T. R. Waller, 2006
- Kaparachlamys Boreham, 1965 †
- Mesopeplum Iredale, 1929
- Phialopecten Marwick, 1928 †
- Sectipecten Marwick, 1928 †
- Towaipecten Beu, 1995 †
- Suku Palliolini Waller, 1993
- Karnekampia H. P. Wagner, 1988
- Lissochlamys Sacco, 1897
- Palliolum Monterosato, 1884
- Placopecten Verrill, 1897
- Pseudamussium Mörch, 1853
- Suku Serripectinini T. R. Waller, 2006 †
- Janupecten Marwick, 1928 †
- Serripecten Marwick, 1928 †
- Suku Adamussiini Habe, 1977
- Subkeluarga Pectininae
- Suku Aequipectinini F. Nordsieck, 1969
- Aequipecten P. Fischer, 1886
- Argopecten Monterosato, 1889
- Cryptopecten Dall, Bartsch & Rehder, 1938
- Flexopecten Sacco, 1897
- Haumea Dall, Bartsch & Rehder, 1938
- Leptopecten Verrill, 1897
- Volachlamys Iredale, 1939
- Suku Amusiini Ridewood, 1903
- Amusium Röding, 1798
- Dentamussium Dijkstra, 1990
- Euvola Dall, 1898
- Leopecten Masuda, 1971
- Ylistrum Mynhardt & Alejandrino, 2014
- Suku Austrochlamydini Jonkers, 2003
- Austrochlamys Jonkers, 2003
- Suku Decatopectinini Waller, 1986
- Anguipecten Dall, Bartsch & Rehder, 1938
- Antillipecten T. R. Waller, 2011 T. R. Waller, 2011
- Bractechlamys Iredale, 1939
- Decatopecten Rüppell in G. B. Sowerby II, 1839
- Excellichlamys Iredale, 1939
- Glorichlamys Dijkstra, 1991
- Gloripallium Iredale, 1939
- Juxtamusium Iredale, 1939
- Lyropecten Conrad, 1862
- Mirapecten Dall, Bartsch & Rehder, 1938
- Nodipecten Dall, 1898
- Suku Pectinini Wilkes, 1810
- Annachlamys Iredale, 1939
- †Fascipecten Freneix, Karache & Salvat 1971
- †Gigantopecten Rovereto, 1899
- Minnivola Iredale, 1939
- †Oopecten Sacco, 1897
- †Oppenheimopecten Teppner, 1922
- Pecten Müller, 1776
- Serratovola Habe, 1951
- Suku Aequipectinini F. Nordsieck, 1969
- Subkeluarga Pedinae Bronn, 1862
- Suku Chlamydini von Teppner, 1922
- Austrohinnites Beu & Darragh, 2001 †
- Azumapecten Habe, 1977
- Chesapecten Ward & Blackwelder, 1975 †
- Chlamys Röding, 1798
- Chokekenia Santelli & del Río, 2018 †
- Ckaraosippur Santelli & del Río, 2019 †
- Complicachlamys Iredale, 1939
- Coralichlamys Iredale, 1939
- Dietotenhosen Santelli & del Río, 2019 †
- Equichlamys Iredale, 1929
- Hemipecten A. Adams & Reeve, 1849
- Hinnites Deference, 1821
- Laevichlamys Waller, 1993
- Manupecten Monterosato, 1872
- Moirechlamys Santelli & del Río, 2018 †
- Notochlamys Cotton, 1930
- Pascahinnites Dijkstra & Raines, 1999
- Pixiechlamys Santelli & del Río, 2018 †
- Praechlamys Allasinaz, 1972 †
- Scaeochlamys Iredale, 1929
- Semipallium Jousseaume in Lamy, 1928
- Swiftopecten Hertlein, 1936
- Talochlamys Iredale, 1929
- Veprichlamys Iredale, 1929
- Yabepecten Masuda, 1963 †
- Zygochlamys Ihering, 1907
- Suku Crassadomini Waller, 1993
- Caribachlamys Waller, 1993
- Crassadoma Bernard, 1986
- Suku Fortipectinini Masuda, 1963
- Fortipecten Yabe & Hatai, 1940 †
- Kotorapecten Masuda, 1962 †
- Masudapecten Akiyama, 1962 †
- Mizuhopecten Masuda, 1963
- Nipponopecten Masuda, 1962 †
- Patinopecten Dall, 1898
- Suku Mimachlamydini Waller, 1993
- Mimachlamys Iredale, 1929
- Spathochlamys Waller, 1993
- Suku Pedini Bronn, 1862
- Pedum Bruguière, 1792
- Suku Chlamydini von Teppner, 1922
- Subkeluarga incertae sedis
- Agerchlamys Damborenea, 1993 †
- Athlopecten Marwick, 1928 †
- Camptochlamys Arkell, 1930 †
- Indopecten Douglas, 1929 †
- Jorgechlamys del Río, 2004 †
- Lamellipecten Dijkstra & Maestrati, 2010
- Lindapecten Petuch, 1995
- Mixtipecten Marwick, 1928 †
- Pseudopecten Bayle, 1878 †

Industri makanan laut
[sunting | sunting sumber]Akuakultur
[sunting | sunting sumber]Akuakultur kapis merupakan aktiviti komersial menternak kapis sehingga ia mencapai saiz yang boleh dipasarkan dan dijual sebagai barangan pengguna. Anak-anak kekapis liar dikumpul untuk diternak di Jepun seawal tahun 1934.[29] Cubaan terawal untuk menternak kekapis sepenuhnya dalam persekitaran ladang hanya tercatat pada 1950-an dan 1960-an.[30] Sebelum ini, pengusaha lebih gemar menangkap kekapis liar kerana penternakan boleh menelan belanja besar. Akan tetapi, kemerosotan kekapis sedunia telah menyebabkan berkembangnya akuakultur. Industri akuakultur kapis sedunia kini mantap dengan jumlah pengeluaran tahunan dilaporkan mencecah 1,200,000 tan metrik[31] daripada kira-kira 12 spesies. Kira-kira 90 peratus daripada penghasilan tersebut datang dari negara China dan Jepun.
Penangkapan liar
[sunting | sunting sumber]Perusahaan penangkapan kekapis liar terbesar adalah bagi kapis laut Atlantik (Placopecten magellanicus) di pesisiran timur laut Amerika Syarikat dan timur Kanada. Kekapis diperoleh dengan menggunakan pukat tunda dasar. Penghasilan kekapis juga tertumpu di Jepun (liar mahupun ternak) dan China (rata-rata kapis teluk Atlantik diternak).[32]:p.661
Di Selat D'Entrecasteaux di selatan Tasmania, kerja memukat diharamkan pada tahun 1969, dan semenjak itu kekapis di kawasan berkenaan hanya ditangkap oleh penyelam.[33] Cubaan menggunakan bubu berlampu untuk mengumpan udang karang dan ketam membawa kepada penemuan bahawa ia juga berkesan menarik kekapis.[34]
Rujukan
[sunting | sunting sumber]- ^ Shumway & Parsons 2011, m/s. 207.
- ^ Shumway & Parsons 2011, m/s. 124.
- ^ Milsom & Rigby 2009, m/s. 62.
- ^ a b c d e f Drew 1906, m/s. 5–6.
- ^ Shumway & Parsons 2011, m/s. 59.
- ^ a b Hautmann, Michael (2010). "The first scallop" (PDF). Paläontologische Zeitschrift. 84 (2): 317–322. doi:10.1007/s12542-009-0041-5. S2CID 84457522.
- ^ Harris, Olivia K.; Kingston, Alexandra C. N.; Wolfe, Caitlin S.; Ghoshroy, Soumitra; Johnsen, Sönke; Speiser, Daniel I. (2019). "Core–shell nanospheres behind the blue eyes of the bay scallop Argopecten irradians". Journal of the Royal Society Interface. 16 (159). doi:10.1098/rsif.2019.0383. PMC 6833330. PMID 31640501.
- ^ Speiser, Daniel I.; Johnsen, Sönke (29 December 2008). "Comparative Morphology of the Concave Mirror Eyes of Scallops (Pectinoidea)". American Malacological Bulletin. 26 (1–2): 27–33. doi:10.4003/006.026.0204. S2CID 11584708.
- ^ Speiser, Daniel I.; Loew, Ellis R.; Johnsen, Sönke (1 February 2011). "Spectral sensitivity of the concave mirror eyes of scallops: potential influences of habitat, self-screening and longitudinal chromatic aberration". Journal of Experimental Biology. 214 (3): 422–431. doi:10.1242/jeb.048108. PMID 21228201.
- ^ "Eyes detect changing movement patterns: queen scallop". asknature.org.
- ^ Land, M F; Fernald, R D (March 1992). "The Evolution of Eyes". Annual Review of Neuroscience. 15 (1): 1–29. doi:10.1146/annurev.ne.15.030192.000245. PMID 1575438.
- ^ Speiser, Daniel I.; Johnsen, Sönke (1 July 2008). "Scallops visually respond to the size and speed of virtual particles". Journal of Experimental Biology. 211 (13): 2066–2070. doi:10.1242/jeb.017038. PMID 18552295.
- ^ Sun, Xiujun; Liu, Zhihong; Wu, Biao; Zhou, Liqing; Wang, Qi; Wu, Wei; Yang, Aiguo (2018). "Differences between fast and slow muscles in scallops revealed through proteomics and transcriptomics". BMC Genomics. 19 (1): 377. doi:10.1186/s12864-018-4770-2. PMC 5963113. PMID 29783952.
Bahan-bahan disalin dari sumber ini yang terdapat di bawah Lesen Antarabangsa Atribusi 4.0 Creative Commons.
- ^ a b Shumway & Parsons 2011, m/s. 689–690.
- ^ Dore 2013, m/s. 152.
- ^ Gosling 2015, m/s. 29.
- ^ Matlins 2001, m/s. 56.
- ^ a b Bloom, S. (1975). "The motile escape response of a sessile prey: a sponge-scallop mutualism". Journal of Experimental Biology and Ecology. 17 (3): 311–321. doi:10.1016/0022-0981(75)90006-4.
- ^ a b Pitcher, C.R.; Butler, A.J. (1987). "Predation by asteroids, escape response, and morphometrics of scallops with epizoic sponges". Journal of Experimental Marine Biology and Ecology. 112 (3): 233–249. doi:10.1016/0022-0981(87)90071-2.
- ^ a b Forester, A.J. (1979). "The association between the sponge Halichondria panicea (Pallas) and scallop Chlamys varia (L.): a commensal protective mutualism". Journal of Experimental Marine Biology and Ecology. 36 (1): 1–10. doi:10.1016/0022-0981(79)90096-0.
- ^ Donovan, D.; Bingham, B.; Farren, H.; Gallardo, R.; Vigilant, V. (2002). "Effects of sponge encrustation on the swimming behaviour energetics and morphometry of the scallop Chlamys hastata" (PDF). Journal of the Marine Biological Association of the United Kingdom. 82 (3): 469–476. doi:10.1017/s0025315402005738. S2CID 56338849.
- ^ a b c Hart, D.R.; Chute, A.S. (2004). "Essential Fish Habitat Source Document: Sea Scallop, Placopecten magellanicus, Life History and Habitat Characteristics" (PDF). NOAA Tech Memo NMFS NE-189.
- ^ "Scallop Aquaculture" (PDF). College of Marine Science, University of South Florida.
- ^ Rice 2012, m/s. 47.
- ^ Waller, T. R. (1993). "The evolution of Chlamys (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific". American Malacological Bulletin. 10 (2): 195–249.
- ^ Harper dll. 2000, m/s. 254.
- ^ Malkowsky, Yaron; Klussmann-Kolb, Annette (May 2012). "Phylogeny and spatio-temporal distribution of European Pectinidae (Mollusca: Bivalvia)". Systematics and Biodiversity. 10 (2): 233–242. doi:10.1080/14772000.2012.676572. S2CID 84085349.
- ^ Habe 1977.
- ^ Kinoshita T (1935) A test for natural spat collection of the Japanese scallop. Report of the Hokkaido Fish Research Station, 273:1-8.
- ^ Shumway SE & Parsons GJ (2006). Scallops: Biology, Ecology and Aquaculture. Elsevier B.V., Amsterdam.
- ^ "Ifremer". wwz.ifremer.fr. Diarkibkan daripada yang asal pada 9 September 2016. Dicapai pada 13 January 2022.
- ^ Shumway & Parsons 2011, m/s. 902.
- ^ Walker, Margaret (1991). "What price Tasmanian scallops? A report of morbidity and mortality associated with the scallop diving season in Tasmania 1990". South Pacific Underwater Medicine Society Journal. 21 (1). Diarkibkan daripada yang asal pada 2013-10-20. Dicapai pada 2013-07-16.CS1 maint: unfit URL (link)
- ^ "Orkney part of 'scallop discos' fishing trial". BBC News. BBC. 2022-06-06. Dicapai pada 2022-06-07.
| Wikimedia Commons mempunyai media berkaitan Kekapis |
| Ikan |  | |
|---|---|---|
| Kerang-kerangan | ||
| Makanan laut lain | ||
| Makanan laut yang diproses | ||
| Hidangan makanan laut | ||
| Bahaya kesihatan | ||
| Khidmat nasihat | ||
| Kebajikan haiwan | ||
| Topik berkaitan | ||