利用者:Crion/Sandbox
※2010年時点の分類(一部の分類群は2011年)
この記事には独自研究が含まれているおそれがあります。 |

真正細菌の分類(しんせいさいきんのぶんるい)では、これまでに発表された真正細菌の属の一覧とその分類を記述する。2010年終了時点で正式に発表されている狭義の細菌(ドメイン:バクテリア)の属をほぼ全て網羅している。古細菌の10倍以上、約1600属を内包する。
真正細菌の分類は門、綱、目、科、属、種が基本で、それ以外にドメイン、界や亜界、亜綱、亜目、亜種などが使われている。分類自体は16S rRNA系統解析の結果に他の遺伝子の系統解析、生化学的性質を加味して行われているが、一部旧来の分類を引きずっている上、分岐パターンも意見が一致しておらず、今後とも大小の変更がありうる。本項目は基本的に『International Journal of Systematic and Evolutionary Microbiology』、次いで『Bergey's Manual of Systematic Bacteriology (2nd Edition)』に従い記述する。
分類と系統樹の概観
[編集]
- (モネラ界/Monera(真正細菌界/Eubacteria))
- 外膜有り・グラム陰性菌
- #アキドバクテリア門 Acidobacteria
- #アクウィフェクス門 Aquificae
- #アルマティモナス門 Armatimonadetes
- #バクテロイデス門 Bacteroidetes
- #カルディセリクム門 Caldiserica
- #クラミジア門 Chlamydiae (ペプチドグリカン欠損)
- #クロロビウム門 Chlorobi
- #クリシオゲネス門 Chrysiogenetes
- #シアノバクテリア門 Cyanobacteria
- #デフェリバクター門 Deferribacteres
- #デイノコックス-テルムス門 Deinococus-Thermus (一部グラム陽性)
- #エルシミクロビウム門 Elusimicrobia
- #フィブロバクター門 Fibrobacteres
- #フソバクテリア門 Fusobacteria
- #ゲマティモナス門 Gemmatimonadetes
- #レンティスファエラ門 Lentisphaerae
- #ニトロスピラ門 Nitrospira
- #プランクトミケス門 Planctomycetes(ペプチドグリカン欠損)
- #プロテオバクテリア門 Proteobacteria
- #スピロヘータ門 Spirochetes
- #シネルギステス門 Synergistetes
- #サーモデスルフォバクテリア門 Thermodesulfobacteria
- #ウェルコミクロビウム門 Verrucomicrobia
- 外膜無し・グラム陽性菌
- #アクチノバクテリア門 Actinobacteria
- #クロロフレクサス門 Chloroflexi(一部グラム陽性)
- #ディクチオグロムス門 Dictyoglomi(外膜様構造あり・グラム陰性)
- #フィルミクテス門 Firmicutes(一部グラム陰性)
- #テネリクテス門 Tenericutes(細胞壁欠損、グラム陰性)
- #テルモトガ門 Thermotogae(外膜様構造あり・グラム陰性)
- 外膜有り・グラム陰性菌
アクウィフェクス門
[編集]
グラム陰性。広い熱水環境より分離される好気または嫌気性の好熱細菌群。一般に水素を酸化して化学合成独立栄養的に増殖する。最高95°C程度で増殖可能な種を含む(真正細菌の中では最も高い)。真正細菌ではあるが例外的にエーテル結合を含む脂質を持つことでも知られる。16S rRNA系統解析では真正細菌の中で最も深い分岐を示し、コル古細菌と共に生命の起源に最も近い生物とされることもある。一方で、より派生的な系統解析例もあり、プロテオバクテリア門に含まれるか近縁とされる場合がある。
アクウィフェクス綱
[編集]- アクウィフェクス綱 Aquificae Reysenbach 2002
- アクウィフェクス目 Aquificales Reysenbach 2002
- アクウィフェクス科 Aquificaceae Reysenbach 2002
- デスルフォバクテリウム科 Desulfurobacteriaceae L'Haridon et al. 2006
- ヒドロゲノテルムス科 Hydrogenothermaceae Eder and Huber 2003
- 科所属未定
- アクウィフェクス目 Aquificales Reysenbach 2002
テルモトガ門
[編集]グラム陰性。熱水環境や油田より分離される嫌気性の好熱菌(最高90℃程度)で、有機物を還元して化学合成従属栄養的に増殖する。桿菌の端に外側に出っ張った鞘のような構造(トガ)を作る特徴がある。アクウィフェクス門と同じくこの門も16S rRNA系統解析では真正細菌の中で深い分岐を示すが、グラム陽性菌の系統に近縁とされる系統解析例も増えてきている。細胞膜はエーテル結合を持つ脂質を含む。
テルモトガ綱
[編集]- テルモトガ綱 Thermotogae Reysenbach 2002
- テルモトガ目 Thermotogales Reysenbach 2002
- テルモトガ科 Thermotogaceae Reysenbach 2002
- Thermotoga - Fervidobacterium - Geotoga - Kosmotoga - Marinitoga - Petrotoga - Thermococcoides - Thermosipho
- テルモトガ科 Thermotogaceae Reysenbach 2002
- テルモトガ目 Thermotogales Reysenbach 2002
サーモデスルフォバクテリア門
[編集]グラム陰性。独立又は混合栄養性の硫酸還元菌。好熱性で、温泉や熱水噴出孔、汚水処理設備などから分離される[2][3]。
サーモデスルフォバクテリア綱
[編集]- サーモデスルフォバクテリア綱 Thermodesulfobacteria Hatchikian et al. 2002
- サーモデスルフォバクテリア目 Thermodesulfobacteriales Hatchikian et al. 2002
- サーモデスルフォバクテリア科 Thermodesulfobacteriaceae Hatchikian et al. 2002
- Thermodesulfobacterium - Caldimicrobium - Thermodesulfatator
- サーモデスルフォバクテリア科 Thermodesulfobacteriaceae Hatchikian et al. 2002
- サーモデスルフォバクテリア目 Thermodesulfobacteriales Hatchikian et al. 2002
デイノコックス-テルムス門
[編集]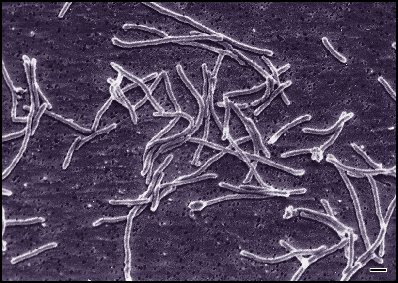
放射線耐性菌デイノコックスと好熱菌であるテルムスの近縁群を中心とした門。従属栄養的で、好気性生物が多い。細胞壁の構造そのものはグラム陰性菌に近く外膜を持つが、非常に厚いペプチドグリカン層を持ちグラム陽性に染まる菌も含まれる。そのため、かつてグラム陽性菌門(フィルミクテス門:当時は現在のフィルミクテス門に加えて放線菌も含んでいた)に分類されたこともある。
系統的には、Guptaらによればグラム陽性菌とグラム陰性菌の中間的な系統になるという。クロロフレクサス門に近縁とも言われる。トーマス・キャバリエ=スミスらによれば、全生物の中でも最も初期に分岐したという。
デイノコックス綱
[編集]- デイノコックス綱 Deinococci Garrity and Holt 2002
- デイノコックス目 Deinococcales Rainey et al. 1997
- デイノコックス科 Deinococcaceae Brooks and Murray 1981
- トルエペラ科 Trueperaceae Rainey, da Costa and Albuquerque 2005
- Truepera
- テルムス目 Thermales Rainey and da Costa 2002
- テルムス科 Thermaceae da Costa and Rainey 2002
- デイノコックス目 Deinococcales Rainey et al. 1997
クリシオゲネス門
[編集]1996年にヒ素で汚染されたヨシ原から単離されたChrysiogenes arsenatis、2007年にオランダの脱硫バイオリアクターより発見されたDesulfurispirillum alkaliphilum2属2種よりなる門。嫌気性で、酸素を利用する代わりに前者はヒ素(ヒ酸塩)、後者は硫黄・硝酸を還元する。グラム陰性短桿菌。
クリシオゲネス綱
[編集]- クリシオゲネス綱 Chrysiogenetes Garrity and Holt 2002
- クリシオゲネス目 Chrysiogenales Garrity and Holt 2002
- クリシオゲネス科 Chrysiogenaceae Garrity and Holt 2002
- Chrysiogenes - Desulfurispira - Desulfurispirillum
- クリシオゲネス科 Chrysiogenaceae Garrity and Holt 2002
- クリシオゲネス目 Chrysiogenales Garrity and Holt 2002
クロロフレクサス門
[編集]酸素非発生型光合成細菌である緑色非硫黄細菌を中心とした門。糸状の群体を形成する好熱性の通性嫌気性菌が多く、温泉などでバクテリアマットを形成している姿が見られることがある。一般に好気条件下では有機物を代謝して活動し、光存在下酸素分圧の低下が起こると光合成系が誘導される。滑走による移動能力を持つ。ただしそれ以外の形質、たとえば光合成系を一切持たず、嫌気条件で有機物を分解して増殖するような種も多い。
グラム陰性菌ではあるが、他のグラム陰性菌とは細胞壁の構造がやや異なっている。好気性の好熱菌、サーモミクロビア綱(#サーモミクロビア門)を含める場合もある。また、シアノバクテリアやデイノコックス・テルムス門に近縁と言われ、これらを含めたテラバクテリア (Terrabacteria)と呼ばれるクレードが提唱されている。
アナエロリネア綱
[編集]- アナエロリネア綱 Anaerolineae Yamada et al. 2006
- アナエロリネア目 Anaerolineales Yamada et al. 2006
- アナエロリネア科 Anaerolineaceae Yamada et al. 2006
- アナエロリネア目 Anaerolineales Yamada et al. 2006
カルディリネア綱
[編集]- カルディリネア綱 Caldilineae Yamada et al. 2006
- カルディリネア目 Caldilineales Yamada et al. 2006
- カルディリネア科 Caldilineaceae Yamada et al. 2006
- カルディリネア目 Caldilineales Yamada et al. 2006
クロロフレクサス綱
[編集]デハロコッコイデス綱
[編集]- デハロコッコイデス綱 "Dehalococcoidetes"
- 目所属未定
- 科所属未定
- Dehalogenimonas
- 科所属未定
- 目所属未定
クテドノバクター綱
[編集]グラム陽性、菌糸や胞子形成など、放線菌に類似する特徴を持つが、系統上はクロロフレクサス門に含まれる。微好気環境で増殖する従属栄養の好熱細菌を幾らか含む[5][6]。
- クテドノバクター綱 Ktedonobacteria corrig. Cavaletti et al. 2007
- クテドノバクター目 Ktedonobacterales corrig. Cavaletti et al. 2007
- クテドノバクター科 Ktedonobacteraceae corrig. Cavaletti et al. 2007
- テルモスポロトリクス科 Thermosporotrichaceae Yabe et al. 2010
- Thermosporothrix Yabe et al. 2010
- クテドノバクター目 Ktedonobacterales corrig. Cavaletti et al. 2007
テルモミクロビウム綱
[編集]1970年代にイエローストーン国立公園から分離されたThermomicrobium roseumと、かつて放線菌に近縁と考えられていたSphaerobacter thermophilus[8]の2属2種のみを含む。独立門とすることもあるが、近年はクロロフレクサス門に含める見解が多い[8]。従属異栄養性の好気性好熱菌で、形状はいびつな桿菌。ピンク色の色素を産生する。
- サーモミクロビア綱 Thermomicrobia Garrity and Holt 2002
- サーモミクロビア目 Thermomicrobiales Garrity and Holt 2002
- サーモミクロビア科 Thermomicrobiaceae Garrity and Holt 2002
- スフェロバクター亜綱 Sphaerobacteridae Stackebrandt et al. 1997
- スフェロバクター目 Sphaerobacterales Stackebrandt et al. 1997
- スフェロバクター亜目 "Sphaerobacterineae"
- スフェロバクター科 Sphaerobacteraceae Stackebrandt et al. 1997
- スフェロバクター亜目 "Sphaerobacterineae"
- スフェロバクター目 Sphaerobacterales Stackebrandt et al. 1997
- サーモミクロビア目 Thermomicrobiales Garrity and Holt 2002
ニトロスピラ門
[編集]グラム陰性のビブリオ様桿菌、またはらせん菌。硝酸菌(Nitrospira)や鉄酸化菌(Leptospirillum)、硫酸還元菌(Thermodesulfovibrio)など独立栄養細菌が多い。
ニトロスピラ綱
[編集]デフェリバクター門
[編集]グラム陰性の直線状またはビブリオ様桿菌。嫌気性の従属栄養細菌が多い。
デフェリバクター綱
[編集]- デフェリバクター綱 Deferribacteres Huber and Stetter 2002
- デフェリバクター目 Deferribacterales Huber and Stetter 2002
- デフェリバクター科 Deferribacteraceae Huber and Stetter 2002
- Deferribacter - Calditerrivibrio - Denitrovibrio - Flexistipes - Geovibrio - Mucispirillum
- 科所属未定
- Caldithrix
- デフェリバクター科 Deferribacteraceae Huber and Stetter 2002
- デフェリバクター目 Deferribacterales Huber and Stetter 2002
シアノバクテリア門
[編集]
光合成によって酸素を放出する細菌。水中に広く分布し、海中、湖沼などに広く見られる。外膜を持ち、グラム染色による分類ではグラム陰性菌に分類される。葉緑体との共通点が多く、一次植物の葉緑体の起源と考えられる。
原核生物としては比較的細胞が大きく、一部は肉眼でも観察可能な糸状・樹状の群体を形成する。真核藻類と大きさや形が比較的類似し、植物と同じく酸素発生型光合成を行うことから、歴史的に植物として扱われてきた経緯を持つ。この時代、藍藻は最も単純で祖先的な植物であると説明された。
しかしその後の研究進展に伴い、藍藻は葉緑体の起源として考えられるようになり、植物との直接的な繋がりは認められなくなった。1970年代から細菌として分類されるべきだと主張され始め、現在では、国際細菌命名規約に従っての記載も行われ、移行期にあるといえる[9]。Bergey’s Manualは、第8版(1974年)からシアノバクテリアを取り上げている。ただし分類は現在でも混乱している。
光合成色素としてフィコシアニンを持つため藍色をしているものが多く、このためギリシャ語の藍色(Κυανος, L:Kyanos)から藍色細菌Cyanobacteria(植物としては藍色植物門; Cyanophyta)と呼ばれる。
- バージェイ式分類における暫定分類
Bergey's Manual of Systematic Bacteriology(2nd Edition, 2001)は、一時的な措置として5つの亜群(Subsection)に分類している。これらは形態に基づく分類であり、自然分類でない可能性が高い。
- シアノバクテリア綱 Cyanobacteria
- 第1亜群/Subsection I(クロオコッカス目 "Chroococcales")
- Family
- Chamaesiphon - Chroococcus - Cyanobacterium - Cyanobium - Cyanothece - Dactylococcopsis - Gloeobacter - Gloeocapsa - Gloeothece - Microcystis - Prochlorococcus - Prochloron - Synechoccus - Synechocystis
- Family
- 第2亜群/Subsection II(プレウロカプサ目/"Pleurocapsales")
- Family
- Cyanocystis - Dermocarpella - Stanieria - Xenococcus
- Family
- Chroococcidiopsis - Myxosarcina - Pleurocapsa-group
- Family
- 第3亜群/Subsection III(ユレモ目/"Oscillatoriales")
- Family
- Arthrospira - Borzia - Crinalium - Geitlerinemia - Leptolyngbya - Limnothrix - Lyngbya - Microcoleus - Oscillatoria - Planktothrix - Prochlorothrix - Pseudanabaena - Spirulina - Starria - Symploca - Trichodesmium - Tychonema
- Family
- 第4亜群/Subsection IV(ネンジュモ目/"Nostocales")
- Family
- Anabaena - Anabaenopsis - Aphanizomenon - Cyanospira - Cylindrospermopsis - Cylindrospermum - Nodularia - Nostoc - Scytonema
- Family
- Calothrix - Rivularia - Tolypothrix
- Family
- 第5亜群/Subsection V(スティゴネマ目/"Stigonematales")
- Family
- Chlorogloeopsis - Fischerella - Geitleria - Iyengariella - Nostochopsis - Stigonema
- Family
- 第1亜群/Subsection I(クロオコッカス目 "Chroococcales")
クロロビウム門
[編集]
偏性嫌気性で緑褐色を呈し、酸素非発生型光合成を行う光合成独立栄養細菌の一群。硫化水素などの硫黄を電子供与体として利用し、光合成により硫黄や硫酸にまで酸化する。酸素の届かない深い水系、泉系に生息する種が多い。
クロロビウム綱
[編集]- クロロビウム綱 "Chlorobia" Chlorobea Cavalier-Smith 2002
- クロロビウム目 Chlorobiales Gibbons and Murray 1978
- クロロビウム科 Chlorobiaceae Copeland 1956
- クロロビウム目 Chlorobiales Gibbons and Murray 1978
イグナウィバクテリウム綱
[編集]- イグナウィバクテリウム綱 Ignavibacteria
- イグナウィバクテリウム目 Ignavibacteriales
- イグナウィバクテリウム科 Ignavibacteriaceae
- Ignavibacterium
- イグナウィバクテリウム科 Ignavibacteriaceae
- イグナウィバクテリウム目 Ignavibacteriales
プロテオバクテリア門
[編集]紅色細菌を中心に16S rRNA配列によって定義された巨大な門。形態や生化学的に多様な種を含み、そのため姿を自在に変えるギリシア神話の神「プロテウス」に因んで命名された。rRNA配列の近縁性に従ってアルファからイプシロンまでの5綱に分けられている。属の数にして真正細菌の4割超、700属以上が含まれる原核生物最大の門、綱の数としてもユリアーキオータ門(古細菌の門の一つ。8綱)に次ぐ多様性がある。グラム染色では陰性。
アルファプロテオバクテリア綱
[編集]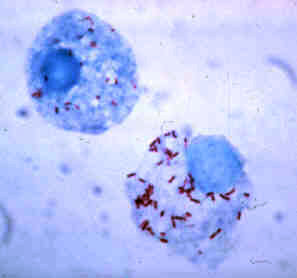
光合成細菌の1種である紅色非硫黄細菌の大部分、例えばロドバクターやロドスピリルムが分類されている。この他に植物や動物の共生体・病原体であるリゾビウムやリケッチア、酢酸発酵菌であるアセトバクターなどが含まれる。ミトコンドリアもこのグループに由来すると考えられている。
- アルファプロテオバクテリア綱 Alphaproteobacteria Garrity et al. 2006
- ロドスピリルム目 Rhodospirillales Pfennig and Trüper 1971
- アセトバクター科 Acetobacteraceae (ex Henrici 1939) Gillis and De Ley 1980
- Acetobacter - Acidicaldus - Acidiphilium - Acidisoma - Acidisphaera - Acidocella - Acidomonas - Ameyamaea - Asaia - Belnapia - Craurococcus - Gluconacetobacter - Gluconobacter - Granulibacter - Kozakia - Neoasaia - Paracraurococcus - Rhodopila - Rhodovarius - Roseococcus - Roseomonas - Rubritepida - Saccharibacter - Stella - Swaminathania - Tanticharoenia - Zavarzinia
- ロドスピリルム科 Rhodospirillaceae Pfennig and Trüper 1971
- Rhodospirillum - Azospirillum - Caenispirillum - Conglomeromonas - Defluviicoccus - Dongia - Fodinicurvata - Inquilinus - Insolitispirillum - Magnetospirillum - Marispirillum - Nisaea - Novispirillum - Oceanibaculum - Pelagibius - Phaeospirillum - Rhodocista - Rhodospira - Rhodovibrio - Roseospira - Skermanella - Telmatospirillum - Thalassobaculum - Thalassospira - Tistlia - Tistrella
- アセトバクター科 Acetobacteraceae (ex Henrici 1939) Gillis and De Ley 1980
- リケッチア目 Rickettsiales Gieszczykiewicz 1939
- アナプラズマ科 Anaplasmataceae Philip 1957
- Anaplasma - Aegyptianella - Cowdria - Ehrlichia - Neorickettsia - Wolbachia - (Xenohaliotis)
- ホロスポラ科 Holosporaceae Görtz and Schmidt 2006
- リケッチア科 Rickettsiaceae Pinkerton 1936
- 科所属未定
- アナプラズマ科 Anaplasmataceae Philip 1957
- ロドバクター目 Rhodobacterales Garrity et al. 2006
- ヒフォモナス科 Hyphomonadaceae Lee et al. 2005
- Hyphomonas - Hellea - Henriciella - Hirschia - Maribaculum - Maricaulis - Oceanicaulis - Ponticaulis - Robiginitomaculum
- ロドバクター科 Rhodobacteraceae Garrity et al. 2006
- Rhodobacter - Agaricicola - Ahrensia - Albimonas - Albidovulum - Amaricoccus - Antarctobacter - Catellibacterium - Celeribacter - Citreicella - Citreimonas - Dinoroseobacter - Donghicola - Gaetbulicola - Gemmobacter - Haematobacter - Hwanghaeicola - Jannaschia - Ketogulonicigenium - Labrenzia - Leisingera - Loktanella - Lutimaribacter - Mameliella - Maribius - Marinovum - Maritimibacter - Marivita - Methylarcula - Nautella - Nereida - Nesiotobacter - Oceanibulbus - Oceanicola - Octadecabacter - Palleronia - Pannonibacter - Paracoccus - Pelagibaca - Pelagicola - Phaeobacter - Pontibaca - Ponticoccus - Pseudorhodobacter - Pseudoruegeria - Pseudovibrio - Rhodobaca - Rhodothalassium - Rhodovulum - Roseibaca - Roseibacterium - Roseibium - Roseicyclus - Roseinatronobacter - Roseisalinus - Roseivivax - Roseobacter - Roseovarius - Rubellimicrobium - Rubribacterium - Rubrimonas - Ruegeria - Sagittula - Salinihabitans - Salipiger - Sediminimonas - Seohaeicola - Shimia - Silicibacter - Staleya - Stappia - Sulfitobacter - Tateyamaria - Thalassobacter - Thalassobius - Thalassococcus - Thioclava - Thiosphaera - Tranquillimonas - Tropicibacter - Tropicimonas - Wenxinia - Woodsholea - Yangia
- ヒフォモナス科 Hyphomonadaceae Lee et al. 2005
- スフィンゴモナス目 Sphingomonadales Yabuuchi and Kosako 2006
- エリスロバクター科 Erythrobacteraceae Lee et al. 2005
- Erythrobacter - Altererythrobacter - Croceicoccus - Erythromicrobium - Porphyrobacter
- スフィンゴモナス科 Sphingomonadaceae Kosako et al. 2000
- Sphingomonas - Blastomonas - Erythromonas - Novosphingobium - Sandaracinobacter - Sandarakinorhabdus - Sphingobium - Sphingopyxis - Sphingosinicella - Stakelama - Zymomonas
- エリスロバクター科 Erythrobacteraceae Lee et al. 2005
- カウロバクター目 Caulobacterales Henrici and Johnson 1935
- カウロバクター科 Caulobacteraceae Henrici and Johnson 1935
- リゾビウム目 Rhizobiales Kuykendall 2006
- アウランチモナス科 "Aurantimonadaceae"
- Aurantimonas - Fulvimarina - Martelella
- バルトネラ科 Bartonellaceae Gieszczykiewicz 1939
- Bartonella - Grahamella - Rochalimaea
- ベイジェリンキア科 Beijerinckiaceae Garrity et al. 2006
- Beijerinckia - Camelimonas - Chelatococcus - Methylocapsa - Methylocella - Methylovirgula
- ブラディリゾビウム科 Bradyrhizobiaceae Garrity et al. 2006
- Bradyrhizobium - Afipia - Agromonas - Balneimonas - Blastobacter - Bosea - Nitrobacter - Oligotropha - Rhodoblastus - Rhodopseudomonas - Salinarimonas
- ブルセラ科 Brucellaceae Breed et al. 1957
- Brucella - Crabtreella - Daeguia - Mycoplana - Ochrobactrum - Paenochrobactrum - Pseudochrobactrum
- コハエシバクテル科 Cohaesibacteraceae Hwang and Cho 2008
- Cohaesibacter
- ヒフォミクロビウム科 Hyphomicrobiaceae Babudieri 1950
- Hyphomicrobium - Ancalomicrobium - Angulomicrobium - Aquabacter - Blastochloris - Cucumibacter - Devosia - Dichotomicrobium - Filomicrobium - Gemmiger - Maritalea - Methylorhabdus - Pedomicrobium - Prosthecomicrobium - Rhodomicrobium - Rhodoplanes - Seliberia - Zhangella
- メチロバクテリウム科 Methylobacteriaceae Garrity et al. 2006
- Methylobacterium - Meganema - Microvirga - Protomonas
- メチロシスティス科 Methylocystaceae Bowman 2006
- Methylocystis - Albibacter - Hansschlegelia - Methylopila - Methylosinus - Pleomorphomonas - Terasakiella
- フィロバクテリウム科 Phyllobacteriaceae Mergaert and Swings 2006
- Phyllobacterium - Aminobacter - Aquamicrobium - Chelativorans - Hoeflea - Mesorhizobium - Nitratireductor - Pseudaminobacter - (Candidatus Liberibacter)
- リゾビウム科 Rhizobiaceae Conn 1938
- Rhizobium("Agrobacterium") - Allorhizobium - Carbophilus - Chelatobacter - Ensifer - Kaistia - Sinorhizobium
- ロドビウム科 Rhodobiaceae Garrity et al. 2006
- Rhodobium - Afifella - Anderseniella - Parvibaculum - Roseospirillum - Tepidamorphus
- キサントバクター科 Xanthobacteraceae Lee et al. 2005
- Xanthobacter - Ancylobacter - Azorhizobium - Labrys - Pseudolabrys - Pseudoxanthobacter - Starkeya
- 科所属未定
- Amorphus - Bauldia - Vasilyevaea
- アウランチモナス科 "Aurantimonadaceae"
- キロニエラ目 Kiloniellales Wiese et al. 2009
- キロニエラ科 Kiloniellaceae Wiese et al. 2009
- Kiloniella
- キロニエラ科 Kiloniellaceae Wiese et al. 2009
- コルディイモナス目 Kordiimonadales Kwon et al. 2005
- パルブルアーキュラ目 "Parvularculales"
- スネアチエラ目 Sneathiellales Kurahashi et al. 2008
- スネアチエラ科 Sneathiellaceae Kurahashi et al. 2008
- 目所属未定
- 科所属未定
- Elioraea - Geminicoccus - Rhizomicrobium
- 科所属未定
- ロドスピリルム目 Rhodospirillales Pfennig and Trüper 1971
ベータプロテオバクテリア綱
[編集]
好気性または通性好気性生物が多く、広範囲の有機物を酸化分解する化学合成従属栄養生物、あるいは無機物を酸化する化学合成独立栄養生物(亜硝酸菌や水素酸化菌、鉄酸化菌など)が含まれている。アルファプロテオバクテリアに含まれなかった、一部の紅色非硫黄細菌(光合成細菌)もこちらに分類されている。
- ベータプロテオバクテリア綱 Betaproteobacteria Garrity et al. 2006
- バークホルデリア目 Burkholderiales Garrity et al. 2006
- バークホルデリア科 Burkholderiaceae Garrity et al. 2006
- Burkholderia - Chitinimonas - Cupriavidus - Lautropia - Limnobacter - Pandoraea - Paucimonas - Polynucleobacter - Ralstonia - Thermothrix - Wautersia
- アルカリゲネス科 Alcaligenaceae De Ley et al. 1986
- Alcaligenes - Achromobacter - Advenella - Azohydromonas - Bordetella - Brackiella - Castellaniella - Derxia - Kerstersia - Oligella - Paenalcaligenes - Paenalcaligenes - Parapusillimonas - Parasutterella - Pelistega - Pigmentiphaga - Pusillimonas - Sutterella - Taylorella
- コマモナス科 Comamonadaceae Willems et al. 1991
- Comamonas - Acidovorax - Albidiferax - Alicycliphilus - Brachymonas - Caenimonas - Caenibacterium - Caldimonas - Curvibacter - Delftia - Diaphorobacter - Giesbergeria - Hydrogenophaga - Hylemonella - Kinneretia - Lampropedia - Limnohabitans - Macromonas - Malikia - Ottowia - Pelomonas - Polaromonas - Pseudacidovorax - Pseudorhodoferax - Ramlibacter - Rhodoferax - Roseateles - Schlegelella - Simplicispira - Tepidicella - Variovorax - Verminephrobacter - Xenophilus
- オキサロバクター科 Oxalobacteraceae Garrity et al. 2006
- Oxalobacter - Collimonas - Duganella - Herbaspirillum - Herminiimonas - Janthinobacterium - Massilia - Naxibacter - Oxalicibacterium - Telluria - Undibacterium
- 科所属未定
- Aquabacterium - Aquincola - Ideonella - Inhella - Leptothrix - Methylibium - Mitsuaria - Paucibacter - Piscinibacter - Rivibacter - Rubrivivax - Sphaerotilus - Tepidimonas - Thiobacter - Thiomonas - Xylophilus
- バークホルデリア科 Burkholderiaceae Garrity et al. 2006
- ヒドロゲノフィルス目 Hydrogenophilales Garrity et al. 2006
- ヒドロゲノフィルス科 Hydrogenophilaceae Garrity et al. 2006
- Hydrogenophilus - Petrobacter - Sulfuricella - Tepidiphilus - Thiobacillus
- ヒドロゲノフィルス科 Hydrogenophilaceae Garrity et al. 2006
- メチロフィルス目 Methylophilales Garrity et al. 2006
- メチロフィルス科 Methylophilaceae Garrity et al. 2006
- Methylophilus - Methylobacillus - Methylotenera - Methylovorus
- メチロフィルス科 Methylophilaceae Garrity et al. 2006
- ナイセリア目 Neisseriales Tønjum 2006
- ナイセリア科 Neisseriaceae Prévot 1933
- Neisseria - Alysiella - Andreprevotia - Aquaspirillum - Aquitalea - Bergeriella - Chitinibacter - Chitinilyticum - Chitiniphilus - Chromobacterium - Conchiformibius - Deefgea - Eikenella - Formivibrio - Gulbenkiania - Iodobacter - Jeongeupia - Kingella - Laribacter - Leeia - Microvirgula - Morococcus - Paludibacterium - Prolinoborus - Pseudogulbenkiania - Silvimonas - Simonsiella - Stenoxybacter - Uruburuella - Vitreoscilla - Vogesella
- ナイセリア科 Neisseriaceae Prévot 1933
- ニトロソモナス目 Nitrosomonadales Garrity et al. 2006
- ニトロソモナス科 Nitrosomonadaceae Garrity et al. 2006
- ガリオネラ科 Gallionellaceae Henrici and Johnson 1935
- スピリリウム科 Spirillaceae Migula 1894
- ロドサイクルス目 Rhodocyclales Garrity et al. 2006
- ロドサイクルス科 Rhodocyclaceae Garrity et al. 2006
- Rhodocyclus - Azoarcus - Azonexus - Azospira - Azovibrio - Dechloromonas - Dechlorosoma - Denitratisoma - Ferribacterium - Methyloversatilis - Propionibacter - Propionivibrio - Quatrionicoccus - Shinella - Sterolibacterium - Thauera - Uliginosibacterium - Zoogloea
- ロドサイクルス科 Rhodocyclaceae Garrity et al. 2006
- プロカバクター目 "Procabacteriales"
- バークホルデリア目 Burkholderiales Garrity et al. 2006
ガンマプロテオバクテリア綱
[編集]



シュードモナス科、ビブリオ科、腸内細菌科など医学的に重要で著名な細菌群が多数含まれている。その他紅色硫黄細菌もこちらに分類されている。
- ガンマプロテオバクテリア綱 Gammaproteobacteria Garrity et al. 2005
- アシドチオバシラス目 Acidithiobacillales Garrity et al. 2005
- アシドチオバシラス科 Acidithiobacillaceae Garrity et al. 2005
- サーミチオバシラス科 Thermithiobacillaceae Garrity et al. 2005
- エロモナス目 Aeromonadales Martin-Carnahan and Joseph 2005
- エロモナス科 Aeromonadaceae Colwell et al. 1986
- Aeromonas - Oceanimonas - Oceanisphaera - Tolumonas - Zobellella
- サクシニビブリオ科 Succinivibrionaceae Hippe et al. 1999
- Succinivibrio - Anaerobiospirillum - Ruminobacter - Succinatimonas - Succinimonas
- エロモナス科 Aeromonadaceae Colwell et al. 1986
- アルテロモナス目 Alteromonadales Bowman and McMeekin 2005
- アルテロモナス科 Alteromonadaceae Ivanova and Mikhailov 2001
- Aestuariibacter - Agarivorans - Aliagarivorans - Alishewanella - Alteromonas - Bowmanella - Glaciecola - Haliea - Marinimicrobium - Marinobacter - Marinobacterium - Melitea - Microbulbifer - Saccharophagus - Salinimonas
- コルウェリア科 Colwelliaceae Ivanova et al. 2004
- フェリモナス科 Ferrimonadaceae Ivanova et al. 2004
- Ferrimonas - Paraferrimonas
- イディオマリナ科 Idiomarinaceae Ivanova et al. 2004
- モリテーラ科 Moritellaceae Ivanova et al. 2004
- Moritella - Paramoritella
- シュードアルテロモナス科 Pseudoalteromonadaceae Ivanova et al. 2004
- Pseudoalteromonas - Algicola
- シクロモナス科 Psychromonadaceae Ivanova et al. 2004
- シェワネラ科 Shewanellaceae Ivanova et al. 2004
- 科所属未定
- アルテロモナス科 Alteromonadaceae Ivanova and Mikhailov 2001
- カルディオバクテリウム目 Cardiobacteriales Garrity et al. 2005
- カルディオバクテリウム科 Cardiobacteriaceae Dewhirst et al. 1990
- クロマチウム目 Chromatiales Imhoff 2005
- クロマチウム科 Chromatiaceae Bavendamm 1924
- Allochromatium - Amoebobacter - Chromatium - Halochromatium - Isochromatium - Lamprobacter - Lamprocystis - Marichromatium - Nitrosococcus - Pfennigia - Rhabdochromatium - Rheinheimera - Thermochromatium - Thioalkalicoccus - Thiobaca - Thiocapsa - Thiococcus - Thiocystis - Thiodictyon - Thioflavicoccus - Thiohalocapsa - Thiolamprovum - Thiopedia - Thiophaeococcus - Thiorhodococcus - Thiorhodovibrio - Thiospirillum
- エクトチオロドスピラ科 Ectothiorhodospiraceae Imhoff 1984
- Alkalilimnicola - Alkalispirillum - Aquisalimonas - Arhodomonas - Ectothiorhodosinus - Ectothiorhodospira - Halorhodospira - Natronocella - Nitrococcus - Thioalkalispira - Thioalkalivibrio - Thiohalospira - Thiorhodospira
- グラヌロシコックス科 Granulosicoccaceae Lee et al. 2008
- Granulosicoccus
- ハロチオバシラス科 Halothiobacillaceae Kelly and Wood 2005
- Halothiobacillus - Thioalkalibacter - Thiofaba - Thiovirga
- クロマチウム科 Chromatiaceae Bavendamm 1924
- エンテロバクター目 "Enterobacteriales"
- 腸内細菌科 Enterobacteriaceae Rahn 1937
- Arsenophonus - Biostraticola - Brenneria - Buchnera - Budvicia - Buttiauxella - Calymmatobacterium - Cedecea - Citrobacter - Cronobacter - Dickeya - Edwardsiella - Enterobacter - Erwinia - Escherichia(タイプ) - Ewingella - Hafnia - Klebsiella - Kluyvera - Leclercia - Leminorella - Levinea - Mangrovibacter - Moellerella - Morganella - Obesumbacterium - Pantoea - Pectobacterium - Photorhabdus - Plesiomonas - Pragia - Proteus - Providencia - Rahnella - Raoultella - Saccharobacter - Salmonella - Samsonia - Serratia - Shigella - Shimwellia - Sodalis - Tatumella - Thorsellia - Trabulsiella - Wigglesworthia - Xenorhabdus - Yersinia - Yokenella
- 腸内細菌科 Enterobacteriaceae Rahn 1937
- レジオネラ目 Legionellales Garrity et al. 2005
- コクシエラ科 Coxiellaceae Garrity et al. 2005
- Coxiella - Aquicella
- レジオネラ科 Legionellaceae Brenner et al. 1979
- Legionella - Fluoribacter - Sarcobium - Tatlockia
- 科所属未定
- コクシエラ科 Coxiellaceae Garrity et al. 2005
- メチロコックス目 Methylococcales Bowman 2005
- クレノトリックス科 Crenotrichaceae Hansgirg 1888
- Crenothrix
- メチロコッカス科 Methylococcaceae Whittenbury and Krieg 1984
- Methylococcus - Methylobacter - Methylocaldum - Methylohalobius - Methylomicrobium - Methylomonas - Methylosarcina - Methylosoma - Methylosphaera - Methylothermus
- クレノトリックス科 Crenotrichaceae Hansgirg 1888
- オセアノスピリルム目 Oceanospirillales Garrity et al. 2005
- アルカニヴォラックス科 Alcanivoracaceae corrig. Golyshin et al. 2005
- Alcanivorax - Fundibacter - Kangiella
- ハヘラ科 Hahellaceae Garrity et al. 2005
- Hahella - Endozoicomonas - Halospina - Kistimonas - Zooshikella
- ハロモナス科 Halomonadaceae Franzmann et al. 1989
- Aidingimonas - Carnimonas - Chromohalobacter - Cobetia - Deleya - Halomonas - Halotalea - Halovibrio - Kushneria - Modicisalibacter - Salinicola - Volcaniella - Zymobacter
- リトリコラ科 Litoricolaceae Kim et al. 2007
- Litoricola
- オセアノスピリルム科 Oceanospirillaceae Garrity et al. 2005
- Amphritea - Balneatrix - Bermanella - Marinomonas - Marinospirillum - Neptuniibacter - Neptunomonas - Nitrincola - Oceaniserpentilla - Oceanobacter - Oceanospirillum - Oleispira - Pseudospirillum - Reinekea - Thalassolituus
- オレイフィラス科 Oleiphilaceae Golyshin et al. 2002
- サッカロスピリルム科 "Saccharospirillaceae"
- 科所属未定
- Salicola - Spongiispira
- アルカニヴォラックス科 Alcanivoracaceae corrig. Golyshin et al. 2005
- パスツレラ目 Pasteurellales Garrity et al. 2005
- パスツレラ科 Pasteurellaceae Pohl 1981
- Actinobacillus - Aggregatibacter - Avibacterium - Basfia - Bibersteinia - Chelonobacter - Gallibacterium - Haemophilus - Histophilus - Lonepinella - Mannheimia - Nicoletella - Pasteurella - Phocoenobacter - Volucribacter
- パスツレラ科 Pasteurellaceae Pohl 1981
- シュードモナス目 Pseudomonadales Orla-Jensen 1921(タイプ)
- モラクセラ科 Moraxellaceae Rossau et al. 1991
- Acinetobacter - Alkanindiges - Branhamella - Enhydrobacter - Moraxella - Perlucidibaca - Psychrobacter
- シュードモナス科 Pseudomonadaceae Winslow et al. 1917
- Azomonas - Azomonotrichon - Azorhizophilus - Azotobacter - Cellvibrio - Chryseomonas - Flavimonas - Mesophilobacter - Pseudomonas - Rhizobacter - Rugamonas - Serpens
- 科所属未定
- Dasania
- モラクセラ科 Moraxellaceae Rossau et al. 1991
- サリニスファエラ目 "Salinisphaerales"
- サリニスファエラ科 "Salinisphaeraceae"
- Salinisphaera
- サリニスファエラ科 "Salinisphaeraceae"
- チオスリックス目 Thiotrichales Garrity et al. 2005
- フランシセラ科 Francisellaceae Sjöstedt 2005
- ピスキリケッチア科 Piscirickettsiaceae Fryer and Lannan 2005
- Cycloclasticus - Hydrogenovibrio - Methylophaga - Piscirickettsia - Sulfurivirga - Thioalkalimicrobium - Thiomicrospira
- チオスリックス科 Thiotrichaceae Garrity et al. 2005
- Achromatium - Beggiatoa - Leucothrix - Thiobacterium - Thiomargarita - Thioploca - Thiospira - Thiothrix
- 科所属未定
- Caedibacter - Fangia
- ビブリオ目 "Vibrionales"
- ビブリオ科 Vibrionaceae Véron 1965
- Aliivibrio - Allomonas - Beneckea - Catenococcus - Enterovibrio - Grimontia - Listonella - Lucibacterium - Photobacterium - Salinivibrio - Vibrio
- ビブリオ科 Vibrionaceae Véron 1965
- キサントモナス目 Xanthomonadales Saddler and Bradbury 2005
- シノバクター科 Sinobacteraceae Zhou et al. 2008
- Alkanibacter - Hydrocarboniphaga - Nevskia - Singularimonas - Sinobacter - Steroidobacter
- キサントモナス科 Xanthomonadaceae Saddler and Bradbury 2005
- Aquimonas - Arenimonas - Dokdonella - Dyella - Frateuria - Fulvimonas - Ignatzschineria - Luteibacter - Luteimonas - Lysobacter - Pseudofulvimonas - Pseudoxanthomonas - Rhodanobacter - Rudaea - Silanimonas - Stenotrophomonas - Thermomonas - Wohlfahrtiimonas - Xanthomonas - Xylella
- シノバクター科 Sinobacteraceae Zhou et al. 2008
- 目所属未定
- 科所属未定
- Alkalimonas - Arenicella - Congregibacter - Gallaecimonas - Gilvimarinus - Marinicella - Methylohalomonas - Methylonatrum - Orbus - Porticoccus - Sedimenticola - Simiduia - Solimonas - Spongiibacter - Thiohalobacter - Thiohalomonas - Thiohalophilus - Thiohalorhabdus - Thioprofundum - Umboniibacter
- 科所属未定
- アシドチオバシラス目 Acidithiobacillales Garrity et al. 2005
デルタプロテオバクテリア綱
[編集]硫酸還元菌と硫黄還元菌がこの綱の主要な構成グループである。この他様々な好気、嫌気性菌が含まれている。この綱に属す粘液細菌 (Myxococcales) は風変りな細菌で、飢餓状態に陥るとあたかも粘菌のように集合し、子実体を形成する。ゲノムサイズが大きく、Sorangium cellulosumは1300万塩基対という出芽酵母を上回るゲノムを持つ。


- デルタプロテオバクテリア綱 Deltaproteobacteria
- デスルフレラ目 Desulfurellales
- デスルフォビブリオ目 Desulfovibrionales
- デスルフォビブリオ科 Desulfovibrionaceae
- Desulfovibrio - Bilophila - Desulfocurvus - Desulfomonas - Lawsonia
- デスルフォミクロビウム科 Desulfomicrobiaceae
- デスルフォハロビウム科 Desulfohalobiaceae
- Desulfohalobium - Desulfonatronospira - Desulfonatronovibrio - Desulfonauticus - Desulfothermus - Desulfovermiculus
- デスルフォナトロヌム科 Desulfonatronaceae
- デスルフォビブリオ科 Desulfovibrionaceae
- デスルフォバクター目 Desulfobacterales
- デスルフォバクター科 Desulfobacteraceae
- Desulfobacter - Desulfatibacillum - Desulfatiferula - Desulfatirhabdium - Desulfobacterium - Desulfobacula - Desulfobotulus - Desulfocella - Desulfococcus - Desulfofaba - Desulfofrigus - Desulfoluna - Desulfomusa - Desulfonema - Desulforegula - Desulfosalsimonas - Desulfosarcina - Desulfospira - Desulfotignum
- デスルフォバルブス科 Desulfobulbaceae
- Desulfobulbus - Desulfocapsa - Desulfofustis - Desulfopila - Desulforhopalus - Desulfotalea - Desulfurivibrio
- ニトロスピナ科 Nitrospinaceae
- デスルフォバクター科 Desulfobacteraceae
- デスルファルクルス目 Desulfarculales
- デスルフロモナス目 Desulfuromonadales
- デスルフロモナス科 Desulfuromonadaceae
- ゲオバクター科 Geobacteraceae
- Geobacter -Geoalkalibacter - Geopsychrobacter - Geothermobacter - Trichlorobacter
- シントロフォバクター目 Syntrophobacterales
- デロビブリオ目 Bdellovibrionales
- デロビブリオ科 Bdellovibrionaceae
- バクテリオヴォラックス科 Bacteriovoracaceae(Peredibacteraceae)
- Bacteriovorax - Bacteriolyticum - Peredibacter
- ミクソコックス目 Myxococcales
- シストバクター亜目 Cystobacterineae
- シストバクター科 Cystobacteraceae
- ミクソコックス科 Myxococcaceae
- Myxococcus - Angiococcus - Corallococcus - Pyxidicoccus
- ソランギウム亜目 Sorangiineae
- ポリアンギウム科 Polyangiaceae
- Polyangium - Byssovorax("Byssophaga") - Chondromyces - Jahnella - Sorangium
- パセリキュシティス科 Phaselicystidaceae
- Phaselicystis
- ポリアンギウム科 Polyangiaceae
- ナンノシスティス亜目 Nannocystineae
- ナンノシスティス科 Nannocystaceae
- Nannocystis - Enhygromyxa - Plesiocystis
- ハリアンギウム科 "Haliangiaceae"
- コフレリア科 Kofleriaceae
- ナンノシスティス科 Nannocystaceae
- シストバクター亜目 Cystobacterineae
- 目所属未定
- シュントロポラブドゥス科 Syntrophorhabdaceae
- Syntrophorhabdus
- シュントロポラブドゥス科 Syntrophorhabdaceae
イプシロンプロテオバクテリア綱
[編集]
らせん菌または歪曲形をした微生物で、殆どがヒトを含む動物の共生体ないし病原体である。ただし、熱水噴出口から見つかったものも存在する。培養が比較的難しく、プロテオバクテリア門の中では最もよく分かっていない。
- イプシロンプロテオバクテリア綱 Epsilonproteobacteria
- カンピロバクター目 Campylobacterales
- カンピロバクター科 Campylobacteraceae
- ヘリコバクター科 Helicobacteraceae
- Helicobacter - Sulfuricurvum - Sulfurimonas - Sulfurovum - Thiovulum - Wolinella
- ヒュドロゲニモナス科 "Hydrogenimonaceae"
- Hydrogenimonas
- ナウチリア目 Nautiliales(カンピロバクター目に含める場合あり)
- ナウチリア科 Nautiliaceae
- Nautilia - Caminibacter - Lebetimonas - Nitratifractor - Nitratiruptor - Thioreductor
- ナウチリア科 Nautiliaceae
- カンピロバクター目 Campylobacterales
ゼータプロテオバクテリア綱
[編集]- ゼータプロテオバクテリア綱 "Zetaproteobacteria"
- マリプロフンドゥス目 "Mariprofundales"
- マリプロフンドゥス科 "Mariprofundaceae"
- Mariprofundus
- マリプロフンドゥス科 "Mariprofundaceae"
- マリプロフンドゥス目 "Mariprofundales"
フィルミクテス門
[編集]グラム陽性でDNA中のGC含量が55%未満(#アクチノバクテリア門は55%以上)の細菌群(ただしグラム陰性に染まる種も散在する)。グラム陽性菌門、低GCグラム陽性菌門などとも呼ばれる。広い範囲に芽胞形成能力を持つ種を含み、この状態になることで熱、乾燥、消毒剤、放射線などに強い耐性を示す。今のところ大まかにいって偏性嫌気性菌がクロストリジウム綱、通性/偏性好気性菌がバシラス綱に分類されているが、それぞれ単系統でないと指摘されており、今後大きな再編がありうる。クロストリジウム綱には光合成細菌も1科(ヘリオバクテリア)含まれている。
細胞壁を欠きグラム陽性に染まらないマイコプラズマ類(モリキューテス綱、モリクテス綱)は塩基配列の相同性によりフィルミクテス門に含まれることがあるが、独立の門とする場合が多い。ネガティウィクテス綱はクロストリジウム綱から分かれる形で2010年新たに設定されたもの、嫌気性でグラム染色ではグラム陰性に染色されるが、外膜を持たないためグラム陽性菌の系統と分かる。この門は16S rRNA系統解析により設置されており、ヘリオバクテリアなど一部のグラム陰性菌はクロストリジウム綱に残留した。
その他、エリシペロトリクス綱(哺乳類や鳥類に分布する通性嫌気性菌)、テルモリトバクテル綱(鉄を還元する偏性嫌気性菌)についても、それぞれ16S rRNA配列の相違により他の綱と区別された。
バシラス綱
[編集]フィルミクテス門のうち、好気性・通性好気性の属が主に分類されている。芽胞を形成するものもしないものもいる。バシラス綱と名づけられているが、球菌も少なくない。




- バシラス綱 Bacilli
- バシラス目 Bacillales
- アリシクロバシラス科 Alicyclobacillaceae
- バシラス科 Bacillaceae
- Aeribacillus - Alkalibacillus - Amphibacillus - Anaerobacillus - Anoxybacillus - Aquisalibacillus - Bacillus - Caldalkalibacillus - Cerasibacillus - Falsibacillus - Filobacillus - Geobacillus - Gracilibacillus - Halalkalibacillus - Halobacillus - Halolactibacillus - Lentibacillus - Lysinibacillus - Marinococcus - Natronobacillus - Oceanobacillus - Ornithinibacillus - Paraliobacillus - Paucisalibacillus - Pelagibacillus - Piscibacillus - Pontibacillus - Saccharococcus - Salibacillus - Salimicrobium - Salinibacillus - Salirhabdus - Salsuginibacillus - Sediminibacillus - Tenuibacillus - Terribacillus - Thalassobacillus - Tumebacillus - Virgibacillus - Viridibacillus - Vulcanibacillus
- リステリア科 Listeriaceae
- パエニバシラス科 Paenibacillaceae
- Ammoniphilus - Aneurinibacillus - Brevibacillus - Cohnella - Fontibacillus - Oxalophagus - Paenibacillus - Saccharibacillus - Thermobacillus
- パステウリア科 Pasteuriaceae
- プラノコッカス科 Planococcaceae
- Bhargavaea - Caryophanon - Filibacter - Jeotgalibacillus - Kurthia - Marinibacillus - Paenisporosarcina - Planococcus - Planomicrobium - Sporosarcina - Ureibacillus
- スポロラクトバシラス科 Sporolactobacillaceae
- Pullulanibacillus - Sinobaca - Sporolactobacillus - Tuberibacillus
- スタフィロコッカス科 Staphylococcaceae
- Jeotgalicoccus - Macrococcus - Nosocomiicoccus - Salinicoccus - Staphylococcus
- サーモアクチノミセス科 Thermoactinomycetaceae
- Desmospora - Laceyella - Mechercharimyces - Planifilum - Seinonella - Shimazuella - Thermoactinomyces - Thermoflavimicrobium
- 科未帰属
- Exiguobacterium - Gemella - Geomicrobium - Rummeliibacillus - Solibacillus - Thermicanus
- ラクトバシラス目 Lactobacillales
- エロコッカス科 Aerococcaceae
- カルノバクテリウム科 Carnobacteriaceae
- Agitococcus - Alkalibacterium -Allofustis - Alloiococcus - Atopobacter - Atopococcus - Atopostipes - Carnobacterium - Desemzia - Dolosigranulum - Granulicatella - Isobaculum - Lacticigenium - Lactosphaera - Marinilactibacillus - Trichococcus
- エンテロコッカス科 Enterococcaceae
- Bavariicoccus - Catellicoccus - Enterococcus - Melissococcus - Pilibacter - Tetragenococcus - Vagococcus
- ラクトバシラス科 Lactobacillaceae
- Lactobacillus - Paralactobacillus - Pediococcus - Sharpea
- ロイコノストック科 "Leuconostocaceae"
- Leuconostoc - Fructobacillus - Oenococcus - Weissella
- ストレプトコッカス科 Streptococcaceae
- Streptococcus - Lactococcus - Lactovum
- バシラス目 Bacillales
クロストリジウム綱
[編集]フィルミクテス門のうち、偏性嫌気性の属の大部分がクロストリジウム綱に分類される。大半の属が芽胞形成能力を持つ。ヘリオバクテリアなどグラム陰性の科も一部含む。


- クロストリジウム綱 Clostridia
- クロストリジウム目 Clostridiales
- カルディコプロバクター科 Caldicoprobacteraceae
- Caldicoprobacter
- クロストリジウム科 Clostridiaceae
- Alkaliphilus - Anaerobacter - Anaerosporobacter - Anoxynatronum - Butyricicoccus - Caloramator - Caloranaerobacter - Caminicella - Clostridiisalibacter - Clostridium - Fervidicella - Geosporobacter - Lactonifactor - Lutispora - Natronincola - Oxobacter - Proteiniclasticum - Saccharofermentans - Sarcina - Sporacetigenium - Tepidimicrobium - Thermobrachium - Thermohalobacter - Thermotalea - Tindallia
- ユウバクテリウム科 Eubacteriaceae
- Acetobacterium - Alkalibacter - Alkalibaculum - Anaerofustis - Eubacterium - Garciella - Pseudoramibacter
- グラシリバクター科 Gracilibacteraceae
- Gracilibacter
- ヘリオバクテリア科 Heliobacteriaceae
- ラクノスピラ科 Lachnospiraceae
- Acetitomaculum - Anaerostipes - Butyrivibrio - Catonella - Cellulosilyticum - Coprococcus - Dorea - Hespellia - Johnsonella - Lachnobacterium - Lachnospira - Marvinbryantia - Moryella - Oribacterium - Parasporobacterium - Pseudobutyrivibrio - Robinsoniella - Roseburia - Shuttleworthia - Sporobacterium - Syntrophococcus
- オシロスピラ科 Oscillospiraceae
- Oscillospira - Oscillibacter
- ペプトコッカス科 Peptococcaceae
- Cryptanaerobacter - Dehalobacter - Desulfitibacter - Desulfitispora - Desulfitobacterium - Desulfonispora - Desulfosporosinus - Desulfotomaculum - Desulfurispora - Pelotomaculum - Peptococcus - Sporotomaculum - Syntrophobotulus - Thermincola - Thermoterrabacterium
- ペプトストレプトコッカス科 Peptostreptococcaceae
- ルミノコッカス科 Ruminococcaceae
- Acetanaerobacterium - Acetivibrio - Anaerofilum - Anaerotruncus - Ethanoligenens - Faecalibacterium - Fastidiosipila - Hydrogenoanaerobacterium - Papillibacter - Ruminococcus - Sporobacter - Subdoligranulum
- シントロフォモナス科 Syntrophomonadaceae
- Dethiobacter - Fervidicola - Pelospora - Syntrophomonas - Syntrophospora - Syntrophothermus - Thermohydrogenium - Thermosyntropha
- 科未帰属
- Acetoanaerobium - Acidaminobacter - Anaerobranca - Anaerococcus - Anaerosphaera - Anaerovirgula - Anaerovorax - Blautia - Carboxydocella - Dethiosulfatibacter - Finegoldia - Flavonifractor - Flavonifractor - Fusibacter - Gallicola - Guggenheimella - Helcococcus - Howardella - Mogibacterium - Murdochiella - Parvimonas - Peptoniphilus - Proteiniborus - Proteocatella - Pseudoflavonifractor - Sedimentibacter - Soehngenia - Sporanaerobacter - Sulfobacillus - Symbiobacterium - Thermaerobacter - Tissierella
- カルディコプロバクター科 Caldicoprobacteraceae
- ハロアナエロビウム目 Halanaerobiales
- ハロアナエロビウム科 Halanaerobiaceae
- Halanaerobium - Halarsenatibacter - Halocella - Haloincola - Halothermothrix
- ハロバクテロイデス科 Halobacteroidaceae
- Acetohalobium - Halanaerobacter - Halobacteroides - Halanaerobaculum - Halonatronum - Natroniella - Orenia - Selenihalanaerobacter - Sporohalobacter
- ハロアナエロビウム科 Halanaerobiaceae
- ナトロアナエロビウス目 Natranaerobiales
- ナトロアナエロビウス科 Natranaerobiaceae
- Natranaerobius - Natronovirga
- ナトロアナエロビウス科 Natranaerobiaceae
- サーモアナエロバクター目 Thermoanaerobacterales
- サーモアナエロバクター科 Thermoanaerobacteraceae
- Acetogenium - Ammonifex - Caldanaerobacter - Caldanaerobius - Carboxydibrachium - Carboxydothermus - Desulfovirgula - Gelria - Moorella - Tepidanaerobacter - Thermacetogenium - Thermanaeromonas - Thermoanaerobacter - Thermoanaerobium - Thermobacteroides
- サーモアデスルフォビウム科 Thermodesulfobiaceae
- Caldanaerovirga - Coprothermobacter - Thermodesulfobium
- 科未帰属
- Caldicellulosiruptor - Mahella - Thermoanaerobacterium - Thermosediminibacter - Thermovenabulum
- サーモアナエロバクター科 Thermoanaerobacteraceae
- クロストリジウム目 Clostridiales
ネガティウィクテス綱
[編集]グラム陰性菌の一群。他のグラム陽性菌と同様、外膜を欠損する。偏性嫌気性。
- ネガティウィクテス綱 Negativicutes[10]
- セレノモナス目 Selenomonadales
- アキダミノコックス科 Acidaminococcaceae
- ウェイッロネッラ科 Veillonellaceae
- Acetonema - Allisonella - Anaeroarcus - Anaeroglobus - Anaeromusa - Anaerosinus - Anaerovibrio - Centipeda - Dendrosporobacter - Dialister - Megamonas - Megasphaera - Mitsuokella - Negativicoccus - Pectinatus - Pelosinus - Propionispira - Propionispora - Quinella - Schwartzia - Selenomonas - Sporolituus - Sporomusa - Sporotalea - Thermosinus - Veillonella - Zymophilus
- セレノモナス目 Selenomonadales
モリクテス綱
[編集]- モリクテス綱 Mollicutes(→#テネリクテス門)
エリシペロトリクス綱
[編集]
通性好気性または偏性嫌気性。芽胞や鞭毛を形成しない。
- エリシペロトリクス綱 Erysipelotrichia
- エリシペロトリクス目 Erysipelotrichales
- エリシペロトリクス科 Erysipelotrichaceae
- Allobaculum - Bulleidia - Catenibacterium - Coprobacillus - Erysipelothrix - Holdemania - Solobacterium - Turicibacter
- エリシペロトリクス科 Erysipelotrichaceae
- エリシペロトリクス目 Erysipelotrichales
テルモリトバクテル綱
[編集]鉄を還元する偏性嫌気性菌Thermolithobacter1属のみ含む。
アクチノバクテリア門
[編集]グラム陽性でDNA中のGC含量が55%以上(おおむね70%付近)の細菌群。いわゆる放線菌(菌糸と胞子を形成し、偏性好気性)を中心に近縁群をまとめたもので、抗生物質を始めとした生理活性物質を合成する種が多数含まれる。アクチノバクテリア門、放線菌門、高GCグラム陽性菌門などと呼ばれる。
トーマス・キャバリエ=スミスを始めとした一部の研究者は、アクチノバクテリアとネオムラ(古細菌+真核生物)が姉妹群であると主張している。実際にアクチノバクテリアは他の真正細菌と異なるいくつかの特徴を持っており、この群から抗生物質耐性と高熱への適応を経たのがネオムラだとしている。しかしながらそれ以外の違いが大きいためこの説は広く受け入れられていない。
アクチノバクテリア綱
[編集]- 放線菌綱 Actinobacteria
- アシディミクロビウム亜綱 Acidimicrobidae
- アシディミクロビウム目 Acidimicrobiales
- アシディミクロビウム亜目 "Acidimicrobineae"
- アシディミクロビウム科 Acidimicrobiaceae
- Acidimicrobium - Ferrimicrobium - Ferrithrix - Ilumatobacter
- イアミア科 Iamiaceae
- Iamia
- アシディミクロビウム科 Acidimicrobiaceae
- アシディミクロビウム亜目 "Acidimicrobineae"
- アシディミクロビウム目 Acidimicrobiales
- ルブロバクター亜綱 Rubrobacteridae
- コリオバクテリウム亜綱 en:Coriobacteridae
- コリオバクテリウム目 Coriobacteriales
- コリオバクテリウム亜目 "Coriobacterineae"
- コリオバクテリウム科 Coriobacteriaceae
- Coriobacterium - Adlercreutzia - Asaccharobacter - Atopobium - Collinsella - Cryptobacterium - Denitrobacterium - Eggerthella - Enterorhabdus - Gordonibacter - Olsenella - Paraeggerthella - Slackia
- コリオバクテリウム科 Coriobacteriaceae
- コリオバクテリウム亜目 "Coriobacterineae"
- コリオバクテリウム目 Coriobacteriales
- ニトリリルプトル亜綱 Nitriliruptoridae
- エウゼビュア目 Euzebyales
- エウゼビュア科 Euzebyaceae
- Euzebya
- エウゼビュア科 Euzebyaceae
- ニトリリルプトル目 Nitriliruptorales
- ニトリリルプトル科 Nitriliruptoraceae
- Nitriliruptor
- ニトリリルプトル科 Nitriliruptoraceae
- エウゼビュア目 Euzebyales
- #アクチノバクテリア亜綱 Actinobacteridae
- アシディミクロビウム亜綱 Acidimicrobidae
アクチノバクテリア亜綱
[編集]- アクチノバクテリア亜綱 Actinobacteridae
- アクチノマイセス目 Actinomycetales
- アクチノマイセス亜目 Actinomycineae
- アクチノマイセス科 Actinomycetaceae
Actinomyces naeslundii Actinomyces israelii
- アクチノマイセス科 Actinomycetaceae
- アクチノポリスポラ亜目 Actinopolysporineae
- マイクロコッカス亜目 Micrococcineae
- マイクロコッカス科 Micrococcaceae
- Micrococcus - Acaricomes - Arthrobacter - Citricoccus - Kocuria - Nesterenkonia - Renibacterium - Rothia - Sinomonas - Stomatococcus - Zhihengliuella
- ラロバクター科 Rarobacteraceae
- ボゴリエラ科 Bogoriellaceae
- ルアニア科 Ruaniaceae
- Haloactinobacterium - Ruania
- サングイバクター科 Sanguibacteraceae
- ブレビバクテリウム科 Brevibacteriaceae
- セルロモナス科 Cellulomonadaceae
- Cellulomonas - Actinotalea - Paraoerskovia - Oerskovia - Tropheryma
- デルマバクター科 Dermabacteraceae
- Dermabacter - Brachybacterium - Devriesea - Helcobacillus
- デルマトフィラス科 Dermatophilaceae
- デルマコッカス科 Dermacoccaceae
- Dermacoccus - Demetria - Kytococcus - Luteipulveratus - Yimella
- イントラスポランギウム科 Intrasporangiaceae
- Intrasporangium - Arsenicicoccus - Fodinibacter - Humibacillus - Humihabitans - Janibacter - Knoellia - Kribbia - Lapillicoccus - Marihabitans - Ornithinicoccus - Ornithinimicrobium - Oryzihumus - Phycicoccus - Serinicoccus - Terrabacter - Terracoccus - Tetrasphaera - ("Candidatus Nostocoida")
- ヨネシア科 Jonesiaceae
- ミクロバクテリウム科 Microbacteriaceae
- Microbacterium - Agreia - Agrococcus - Agromyces - Aureobacterium - Chryseoglobus - Clavibacter - Cryobacterium - Curtobacterium - Frigoribacterium - Frondihabitans - Glaciibacter - Gulosibacter - Humibacter - Klugiella - Labedella - Leifsonia - Leucobacter - Marisediminicola - Microcella - Microterricola - Mycetocola - Okibacterium - Phycicola - Plantibacter - Pseudoclavibacter - Rathayibacter - Rhodoglobus - Salinibacterium - Schumannella - Subtercola - Yonghaparkia - Zimmermannella
- エウテンベルギア科 Beutenbergiaceae
- Beutenbergia - Salana - Serinibacter
- プロミクロモノスポラ科 Promicromonosporaceae
- Promicromonospora - Cellulosimicrobium - Isoptericola - Myceligenerans - Plantactinospora - Xylanibacterium - Xylanimonas
- ヤニエラ科 Yaniellaceae
- Yaniella
- 科所属未定
- Demequina - Koreibacter - Luteimicrobium
- マイクロコッカス科 Micrococcaceae
- コリネバクテリウム亜目 Corynebacterineae
- コリネバクテリウム科 Corynebacteriaceae
- Corynebacterium - Bacterionema - Caseobacter - Turicella
- デイエッチア科 Dietziaceae
- マイコバクテリウム科 Mycobacteriaceae
- Mycobacterium - Amycolicicoccus
- ノカルディア科 Nocardiaceae
- Gordonia - Micropolyspora - Millisia - Nocardia - Rhodococcus - Skermania - Smaragdicoccus - Williamsia
- ゴルドニア科/Gordoniaceae(→ノカルディア科に統合)
- ツカムレラ科 Tsukamurellaceae
- セグニリパラス科 Segniliparaceae
- Segniliparus
- 所属未定
- Hoyosella - Tomitella
- コリネバクテリウム科 Corynebacteriaceae
- ミクロモノスポラ亜目 Micromonosporineae
- ミクロモノスポラ科 Micromonosporaceae
- Micromonospora - Phytohabitans - Actinocatenispora - Actinoplanes - Amorphosporangium - Ampullariella - Asanoa - Catellatospora - Catelliglobosispora - Catenuloplanes - Couchioplanes - Dactylosporangium - Hamadaea - Krasilnikovia - Longispora - Luedemannella - Pilimelia - Planopolyspora - Planosporangium - Polymorphospora - Pseudosporangium - Rugosimonospora - Salinispora - Spirilliplanes - Verrucosispora - Virgisporangium
- ミクロモノスポラ科 Micromonosporaceae
- プロピオニバクテリウム亜目 Propionibacterineae
- プロピオニバクテリウム科 Propionibacteriaceae
- Propionibacterium - Aestuariimicrobium - Arachnia - Brooklawnia - Granulicoccus - Jiangella - Luteococcus - Microlunatus - Propioniferax - Propionimicrobium - Tessaracoccus
- ノカルジオイデス科 Nocardioidaceae
- Nocardioides - Actinopolymorpha - Aeromicrobium - Friedmanniella - Hongia - Kribbella - Marmoricola - Micropruina - Pimelobacter - Propionicicella - Propionicimonas
- プロピオニバクテリウム科 Propionibacteriaceae
- シュードノカルジア亜目 Pseudonocardineae
- シュードノカルジア科 Pseudonocardiaceae
- Pseudonocardia - Actinoalloteichus - Actinobispora - Actinomycetospora - Actinophytocola - Allokutzneria - Amycolata - Amycolatopsis - Crossiella - Faenia - Goodfellowiella - Kibdelosporangium - Kutzneria - Prauserella - Pseudoamycolata - Saccharomonospora - Saccharopolyspora - Sciscionella - Streptoalloteichus - Thermobispora - Thermocrispum
- アクチノシンネーマ科 Actinosynnemataceae
- Actinosynnema - Actinokineospora - Alloactinosynnema - Lechevalieria - Lentzea - Saccharothrix - Umezawaea
- 科所属不明
- Haloechinothrix
- シュードノカルジア科 Pseudonocardiaceae
- ストレプトマイセス亜目 Streptomycineae
- ストレプトマイセス科 Streptomycetaceae
Streptomyces sp. - Streptomyces - Actinopycnidium - Actinosporangium - Chainia - Elytrosporangium - Kitasatoa - Kitasatospora - Microellobosporia - Streptacidiphilus - Streptoverticillium
- ストレプトマイセス科 Streptomycetaceae
- ストレプトスポランギウム亜目 Streptosporangineae
- ストレプトスポランギウム科 Streptosporangiaceae
- Streptosporangium - Acrocarpospora - Herbidospora - Microbispora - Microtetraspora - Nonomuraea - Planobispora - Planomonospora - Planotetraspora - Sphaerisporangium - Thermopolyspora
- ノカルジオプシス科 Nocardiopsaceae
- Nocardiopsis - Haloactinospora - Marinactinospora - Murinocardiopsis - Streptomonospora - Thermobifida
- サーモモノスポラ科 Thermomonosporaceae
- Thermomonospora - Actinoallomurus - Actinocorallia - Actinomadura - Excellospora - Spirillospora
- ストレプトスポランギウム科 Streptosporangiaceae
- フランキア亜目 Frankineae
- フランキア科 Frankiaceae
- クリプトスポランギウム科 Cryptosporangiaceae
- ゲオデルマトフィラス科 Geodermatophilaceae
- ミクロスファエラ科 Microsphaeraceae
- スポリキシア科 Sporichthyaceae
- アシドテルムス科 Acidothermaceae
- ナカムラエラ科 Nakamurellaceae
- Humicoccus - Nakamurella - Saxeibacter
- 科所属未定
- Fodinicola
- グリコミセス亜目 Glycomycineae
- グリコミセス科 Glycomycetaceae
- Glycomyces - Haloglycomyces - Stackebrandtia
- グリコミセス科 Glycomycetaceae
- カテニュリスポラ亜目 Catenulisporineae
- アクチノスピカ科 Actinospicaceae
- Actinospica
- カテニュリスポラ科 Catenulisporaceae
- Catenulispora
- アクチノスピカ科 Actinospicaceae
- キネオスポリア亜目 Kineosporiineae
- キネオスポリア科 Kineosporiaceae
- Kineosporia - Angustibacter - Kineococcus - Quadrisphaera
- キネオスポリア科 Kineosporiaceae
- アクチノマイセス亜目 Actinomycineae
- ビフィドバクテリウム目 Bifidobacteriales
Bifidobacterium adolescentis(ビフィズス菌の一種) - 亜目なし
- ビフィドバクテリウム科 Bifidobacteriaceae
- Bifidobacterium - Aeriscardovia - Alloscardovia - Gardnerella - Metascardovia - Parascardovia - Scardovia
- ビフィドバクテリウム科 Bifidobacteriaceae
- 亜目なし
- アクチノマイセス目 Actinomycetales




プランクトミケス門
[編集]複雑な形態を持つ好気性細菌。広い水系に生息する。ペプチドグリカンを欠くこと、発芽により増殖すること、細胞内に核膜様構造を持つなど特異な形質を持つ。未記載ではあるが、嫌気的にアンモニアを酸化する種が含まれる。
プランクトミケス綱
[編集]- プランクトミケス綱 Planctomycetacia
- プランクトミケス目 Planctomycetales
- プランクトミケス科 Planctomycetaceae
- Planctomyces - Blastopirellula - Gemmata - Isosphaera - Pirellula - Rhodopirellula - Schlesneria - Singulisphaera - Zavarzinella
- プランクトミケス科 Planctomycetaceae
- ブロカディア目 "Brocadiales"
- ブロカディア科 "Brocadiaceae"
- "Ca. Brocadia" - "Ca. Kuenenia" - "Ca. Scalindua" - "Ca. Anammoxoglobus" - "Ca. Jettenia"
- ブロカディア科 "Brocadiaceae"
- プランクトミケス目 Planctomycetales
フィシスファエラ綱
[編集]- フィシスファエラ綱 Phycisphaerae
- フィシスファエラ目 Phycisphaerales
- フィシスファエラ科 Phycisphaeraceae
- Phycisphaera
- フィシスファエラ科 Phycisphaeraceae
- フィシスファエラ目 Phycisphaerales
クラミジア門
[編集]
動物や原生生物の細胞内に共生する特異な細菌群。細胞が非常に小さいうえ、独自のエネルギー生産系を持たず、真核生物の細胞内で増殖する。人に対して病原性を示す
クラミジア綱
[編集]スピロヘータ門
[編集]
らせん形をした細長い形態が特徴の細菌群。エンベロープ(被膜)内に鞭毛を持ち、菌体そのものをくねらせて移動する。広い水系に分布するほか、一部は昆虫の共生生物、人の病原体としても存在する。
スピロヘータ綱
[編集]- スピロヘータ綱 Spirochaetes
- スピロヘータ目 Spirochaetales
- ブレビネーマ科 "Brevinemataceae"
- レプトスピラ科 Leptospiraceae
- Leptonema - Leptospira - Turneriella
- セルプリナ科 "Serpulinaceae"-"Brachyspiraceae"
- スピロヘータ科 Spirochaetaceae
- Spirochaeta - Borrelia - Clevelandina - Cristispira - Diplocalyx - Hollandina - Pillotina - Treponema
- 科所属未定
- Exilispira
- スピロヘータ目 Spirochaetales
フィブロバクター門
[編集]主に反芻動物の反芻胃(他の哺乳類にも存在する)でセルロース分解を担っているFibrobacter1属2種のみを含む。偏性嫌気性で、セルロースを分解してコハク酸と酪酸、酢酸、ギ酸などを生成する[11]。
フィブロバクター綱
[編集]アキドバクテリア門
[編集]
難培養性で、好酸菌や水生の動物内を中心に7属8種が分離されているのみ。しかしながら広い環境、特に土壌から相当数の16S rRNA遺伝子クローンが検出されており、かなり大型の門になる可能性がある[12][13][14][15]。
アキドバクテリア綱
[編集]- アキドバクテリア綱 Acidobacteria
- アキドバクテリア目 Acidobacteriales
- アキドバクテリア科 "Acidobacteriaceae"
- Acidobacterium - Edaphobacter - Granulicella - Terriglobus
- アキドバクテリア科 "Acidobacteriaceae"
- アキドバクテリア目 Acidobacteriales
ホロファガ綱
[編集]綱所属未定
[編集]- 綱所属未定 (Subdivision3)
- Bryobacter
バクテロイデス門
[編集]グラム陰性菌。細胞膜にスフィンゴ脂質を含む。ほぼすべての種が従属栄養性。桿菌またはらせん菌で、多くは鞭毛を持たず、滑走により運動するか不動性の種が多い(ただし鞭毛を持つ種もいないわけではない)。人に対して病原性を示す種は少ないが、日和見感染症の原因となることがある。16S rRNA系統解析により以下の3綱に分けられている。
バクテロイデス綱は偏性嫌気性で、腸内細菌として重要。人腸内ではフィルミクテス門と並び最優位のグループである。フラボバクテリア綱、スフィンゴバクテリア綱は偏性または通性好気性で、海洋、土壌、廃水処理系に広く分布する。クロロビウムやフィブロバクター門に近縁で、スフィンゴバクテリア (Sphingobacteria)と呼ばれるクレードに含まれる。
バクテロイデス綱
[編集]
- バクテロイデス綱 "Bacteroidia"
- "バクテロイデス目 Bacteroidales"
- バクテロイデス科 Bacteroidaceae
- Acetofilamentum - Acetomicrobium - Acetothermus - Anaerorhabdus - Bacteroides - Capsularis
- マリニラビリア科 "Marinilabiaceae"
- Alkaliflexus - Anaerophaga - Marinilabilia
- ポルフィロモナス科 "Porphyromonadaceae"
- Barnesiella - Butyricimonas - Dysgonomonas - Odoribacter - Oribaculum - Paludibacter - Parabacteroides - Petrimonas - Porphyromonas - Proteiniphilum - Tannerella
- プレボテラ科 "Prevotellaceae"
- Hallella - Paraprevotella - Prevotella - Xylanibacter
- リケネーラ科 "Rikenellaceae"
- Rikenella - Alistipes
- バクテロイデス科 Bacteroidaceae
- 目所属未定
- 科所属未定
- Phocaeicola
- 科所属未定
- "バクテロイデス目 Bacteroidales"
フラボバクテリア綱
[編集]- フラボバクテリア綱 "Flavobacteria"
- フラボバクテリウム目 "Flavobacteriales"
- ブラッタバクテリウム科 "Blattabacteriaceae"
- クライオモルファ科 Cryomorphaceae
- Brumimicrobium - Crocinitomix - Cryomorpha - Fluviicola - Lishizhenia - Owenweeksia - Sediminitomix - Wandonia
- フラボバクテリウム科 Flavobacteriaceae
- Actibacter - Aequorivita - Aequorivita - Aestuariicola - Algibacter - Aquimarina - Arenibacter - Bergeyella - Bizionia - Capnocytophaga - Cellulophaga - Chryseobacterium - Cloacibacterium - Coenonia - Costertonia - Croceibacter - Croceitalea - Dokdonia - Donghaeana - Elizabethkingia - Empedobacter - Epilithonimonas - Eudoraea - Euzebyella - Flagellimonas - Flaviramulus - Flavobacterium - Formosa - Fulvibacter - Gaetbulibacter - Gaetbulimicrobium - Galbibacter - Gelidibacter - Gillisia - Gilvibacter - Gramella - Hyunsoonleella - Jejuia - Joostella - Kaistella - Kordia - Kriegella - Krokinobacter - Lacinutrix - Leeuwenhoekiella - Leptobacterium - Lutaonella - Lutibacter - Lutimonas - Maribacter - Mariniflexile - Maritimimonas - Meridianimaribacter - Marixanthomonas - Meridianimaribacter - Mesoflavibacter - Mesonia - Muricauda - Muriicola - Myroides - Nonlabens - Olleya - Ornithobacterium - Persicivirga - Pibocella - Planobacterium - Polaribacter - Pseudozobellia - Psychroflexus - Psychroserpens - Riemerella - Robiginitalea - Salinimicrobium - Salegentibacter - Sandarakinotalea - Sediminibacter - Sediminicola - Soonwooa - Stanierella - Stenothermobacter - Subsaxibacter - Subsaximicrobium - Tamlana - Tenacibaculum - Ulvibacter - Vitellibacter - Wautersiella - Weeksella - Winogradskyella - Yeosuana - Zeaxanthinibacter - Zhouia - Zobellia - Zunongwangia
- フラボバクテリウム目 "Flavobacteriales"
スフィンゴバクテリア綱
[編集]- スフィンゴバクテリア綱 "Sphingobacteria"
- スフィンゴバクテリア目 "Sphingobacteriales"
- キチノファガ科 "Chitinophagaceae"
- Balneola - Chitinophaga - Ferruginibacter - Filimonas - Flavihumibacter - Flavisolibacter - Gracilimonas - Lacibacter - Niabella - Niastella - Parasegetibacter - Sediminibacterium - Segetibacter - Terrimonas
- シクロバクテリウム科 "Cyclobacteriaceae"
- Algoriphagus - Aquiflexum - Belliella - Chimaereicella - Cyclobacterium - Echinicola - Fontibacter - Hongiella - Indibacter - Nitritalea
- サイトファーガ科 Cytophagaceae
- Adhaeribacter - Arcicella - Cytophaga - Dyadobacter - Ekhidna - Emticicia - Flectobacillus - Flexibacter - Hymenobacter - Larkinella - Leadbetterella - Litoribacter - Meniscus - Microscilla - Persicitalea - Pontibacter - Rhodocytophaga - Rhodonellum - Rudanella - Runella - Siphonobacter - Spirosoma - Sporocytophaga
- フラモヴァーガ科 "Flammeovirgaceae"
- Fabibacter - Flammeovirga - Flexithrix - Limibacter - Marinicola - Marinoscillum - Marivirga - Perexilibacter - Persicobacter - Rapidithrix - Reichenbachiella - Roseivirga - Thermonema
- ロドテルムス科 "Rhodothermaceae"
- Rhodothermus - Salinibacter - Salisaeta
- サプロスピラ科 "Saprospiraceae"
- Aureispira - Haliscomenobacter - Lewinella - Saprospira
- スフィンゴバクテリウム科 Sphingobacteriaceae
- Mucilaginibacter - Nubsella - Olivibacter - Parapedobacter - Pedobacter - Pseudosphingobacterium - Solitalea - Sphingobacterium
- キチノファガ科 "Chitinophagaceae"
- スフィンゴバクテリア目 "Sphingobacteriales"
不明な系統
[編集]- 綱所属未定
- 目所属未定
- 科所属未定
- Fulvivirga - Marinifilum - Prolixibacter - Toxothrix
- 科所属未定
- 目所属未定
フソバクテリア門
[編集]
嫌気性の桿菌で、動物の消化器官などに存在。人に対して病原性を持つ種も存在する。
フソバクテリア綱
[編集]- フソバクテリア綱 "Fusobacteria"
- フソバクテリア目 "Fusobacteriales"
- フソバクテリア科 "Fusobacteriaceae"
- Fusobacterium - Cetobacterium - Ilyobacter - Propionigenium - Psychrilyobacter
- レプトトリキア科 "Leptotrichiaceae"
- フソバクテリア科 "Fusobacteriaceae"
- フソバクテリア目 "Fusobacteriales"
ウェルコミクロビウム門
[編集]土壌および多様な水系、動物や原生生物の共生体として発見される。存在量は多いと考えられるが、培養不可能な種が多くまだよく分かっていない[16]。系統的にはレンチスファエラ門やクラミジア門に近接すると考えられている。
ウェルコミクロビウム綱
[編集]- ウェルコミクロビウム綱 Verrucomicrobiae
- ウェルコミクロビウム目 Verrucomicrobiales
- ウェルコミクロビウム科 Verrucomicrobiaceae
- Verrucomicrobium - Akkermansia - Haloferula - Luteolibacter - Persicirhabdus - Prosthecobacter - Roseibacillus
- キシフィネマトバクター科 "Xiphinematobacteriaceae"
- ルブリタレア科 "Rubritaleaceae"
- Rubritalea
- ウェルコミクロビウム科 Verrucomicrobiaceae
- ウェルコミクロビウム目 Verrucomicrobiales
オピタタス綱
[編集]- オピタタス綱 Opitutae
- オピタタス目 Opitutales
- プニセイコックス目 Puniceicoccales
- プニセイコックス科 Puniceicoccaceae
- Puniceicoccus - Cerasicoccus - Coraliomargarita - Pelagicoccus
- プニセイコックス科 Puniceicoccaceae
ディクチオグロムス門
[編集]2008年現在、熱水泉と板紙工場で発見された嫌気性好熱菌であるDictyoglomusの1属2種のみが知られている。植物の細胞壁構成成分であるキシランの資化能を持つ。
ディクチオグロムス綱
[編集]ゲマティモナス門
[編集]リン除去用活性汚泥システムから発見されたGemmatimonas aurantiacaの1属1種のみ。ポリリン酸を蓄積する。
ゲマティモナス綱
[編集]レンティスファエラ門
[編集]海岸、糞便などから分離。培養不可能な種が多く、まだよく分かっていない。海洋などに広がると考えられる。
レンティスファエラ綱
[編集]シネルギステス門
[編集]グラム陰性桿菌、嫌気性。動物の消化器官や嫌気性の廃水処理設備などから見つかるグループ。一部の種は動物の感染症に関連するとみられる。何れも偏性嫌気性菌で、アミノ酸、ペプチドまたは、脂肪酸の発酵を行う(一部の種は硫黄を還元する)。
シネルギステス綱
[編集]- シネルギステス綱 Synergistia
- シネルギステス目 Synergistales
- シネルギステス科 Synergistaceae
- Synergistes - Aminobacterium - Aminomonas - Anaerobaculum - Cloacibacillus - Dethiosulfovibrio - Jonquetella - Pyramidobacter - Thermovirga - Thermanaerovibrio
- シネルギステス科 Synergistaceae
- シネルギステス目 Synergistales
テネリクテス門
[編集]
マイコプラズマ及び近縁な細菌より成る分類群である。真核生物(動物や植物)の寄生生物として存在し、多くの場合寄生生活のためステロールなどの複雑な栄養要求性を示す。ゲノムサイズが小さく0.58-1.4Mbp、GC含率も低いことが特徴で、18-40%程度。何れもかなり極端な値を示している。ペプチドグリカン細胞壁を欠くためグラム染色できないが、外膜を所持しておらず、系統的にもグラム陽性菌、特にフィルミクテス門に近縁という。#フィルミクテス門に含める場合もある。
モリクテス綱
[編集]- モリクテス綱 Mollicutes
カルディセリクム門
[編集]2007年に小谷温泉より発見。未培養だったOP5と呼ばれる系統に相当するとみられる。嫌気好熱グラム陰性の従属栄養細菌[23]。
カルディセリクム綱
[編集]- カルディセリクム綱 Caldisericia Mori et al. 2009
- カルディセリクム目 Caldisericales Mori et al. 2009
- カルディセリクム科 Caldisericaceae Mori et al. 2009
- カルディセリクム目 Caldisericales Mori et al. 2009
エルシミクロビウム門
[編集]グラム陰性、嫌気従属栄養性。2008年にPachnoda ephippiataと呼ばれるハナムグリの幼虫から報告されたElusimicrobium minutum1属1種のみを含む。この系統を含むグループは、最初シロアリ後腸から発見されたため、Termite group 1、あるいはTG1門と呼ばれることもある。既にゲノム解析も2例行われた[25][26]。
エルシミクロビウム綱
[編集]- エルシミクロビウム綱 Elusimicrobia Geissinger et al. 2010
- エルシミクロビウム目 Elusimicrobiales Geissinger et al. 2010
- エルシミクロビウム科 Elusimicrobiaceae Geissinger et al. 2010
- エルシミクロビウム目 Elusimicrobiales Geissinger et al. 2010
アルマティモナス門
[編集]グラム陰性、好気従属栄養性。2011年に提唱されたもので、かつて難未培養細菌と呼ばれたOP10系統とみられる。ヨシの根圏、火山性土壌などから分離されている。[27][28]
アルマティモナス綱
[編集]- アルマティモナス綱 Armatimonadia Tamaki et al. 2011
- アルマティモナス目 Armatimonadales Tamaki et al. 2011
- アルマティモナス科 Armatimonadaceae Tamaki et al. 2011
- アルマティモナス目 Armatimonadales Tamaki et al. 2011
クトノモナス綱
[編集]- クトノモナス綱 Chthonomonadetes Lee et al. 2011
- クトノモナス目 Chthonomonadales Lee et al. 2011
- クトノモナス科 Chthonomonadaceae Lee et al. 2011
- クトノモナス目 Chthonomonadales Lee et al. 2011
関連項目
[編集]参考文献
[編集]- Cavalier-Smith, T. (2002). “The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification”. Int J Syst Evol Microbiol 52 (Pt 1): 7–76. PMID 11837318.
- Euzéby, J. P. (1998-2008). “LPSN - List of Prokaryotic names with Standing in Nomenclature” (仏語/英語). 2008年12月20日閲覧。
- Gupta, R. S. (2006年). “Phylogeny of Bacteria- Is it a Tangled Web or Can this be Reliably Resolved?” (英語). 2008年12月20日閲覧。
- Garrity, G. M., et al. (2001, 05年) (英語). Bergey's Manual of Systematic Bacteriology, 2nd Edn. Vols. 1, 2.. Springer-Verlag. ISBN 978-0387987712/ISBN 978-0387950402
- 井上勲『藻類30億年の自然史(第2版) - 藻類から見る生物進化・地球・環境』東海大学出版会、2007年。ISBN 978-4-486-01777-6。
- ^ Bacterial/Prokaryotic Phylogeny Webpage
- ^ Jeanthon, C., L'Haridon, S., Cueff, V., Banta, A., Reysenbach, A. L., Prieur, D. (2002). “Thermodesulfobacterium hydrogeniphilum sp. nov., a thermophilic, chemolithoautotrophic, sulfate-reducing bacterium isolated from a deep-sea hydrothermal vent at Guaymas Basin, and emendation of the genus Thermodesulfobacterium”. Int J Syst Evol Microbiol 52: 765-772. doi:10.1099/ijs.0.02025-0. PMID 12054236.
- ^ Moussard, H., L'Haridon, S., Tindall, B. J., Banta, A., Schumann, P., Stackebrandt, E., Reysenbach, A. L., Jeanthon, C. (2004). “Thermodesulfatator indicus gen. nov., sp. nov., a novel thermophilic chemolithoautotrophic sulfate-reducing bacterium isolated from the Central Indian Ridge”. Int J Syst Evol Microbiol 54: 227-233. エラー: 不正なDOI指定です. PMID 14742485.
- ^ Macy, J. M., Nunan, K., Hagen, K. D., Dixon, D. R., Harbour, P. J., Cahill, M., Sly, L. I. (1996). “Chrysiogenes arsenatis gen. nov., sp. nov., a new arsenate-respiring bacterium isolated from gold mine wastewater”. Int J Syst Evol Microbiol 46 (4): 1153-7. エラー: 不正なDOI指定です. PMID 8863450.
- ^ Cavaletti, L., Monciardini, P., Bamonte, R., Schumann, P., Rohde, M., Sosio, M., Donadio, S. (2006). “New Lineage of Filamentous, Spore-Forming, Gram-Positive Bacteria from Soil”. Applied and Environmental Microbiology 72 (6): 4360-9. doi:10.1128/AEM.00132-06. PMID 16751552.
- ^ Yabe, S., Aiba, Y., Sakai, Y., Hazaka, M., Yokota, A. (2010). “Thermosporothrix hazakensis gen. nov., sp. nov., isolated from compost, description of Thermosporotrichaceae fam. nov. within the class Ktedonobacteria Cavaletti et al. 2007 and emended description of the class Ktedonobacteria”. Int. J. Syst. Evol. Microbiol. 60: 1794-1801. doi:10.1099/ijs.0.018069-0. PMID 19767365.
- ^ Jackson, T. J., R. F. Ramaley, and W. G. Meinschein (1973). “Thermomicrobium, a New Genus of Extremely Thermophilic Bacteria”. Int J Syst Bacteriol 23: 28-36. エラー: 不正なDOI指定です.
- ^ a b Hugenholtz, P., Stackebrandt, E. (2004). “Reclassification of Sphaerobacter thermophilus from the subclass Sphaerobacteridae in the phylum Actinobacteria to the class Thermomicrobia (emended description) in the phylum Chloroflexi (emended description)”. Int J Syst Bacteriol 54: 2049-51. doi:10.1099/ijs.0.03028-0. PMID 15545432.
- ^ Ahoren Oren (2004). “A proposal for further integration of the cyanobacteria under the Bacteriological Code”. Int. J. Syst. Evol. Microbiol. 54: 1895–1902. doi:10.1099/ijs.0.03008-0. PMID 15388760.
- ^ Marchandin, H., Teyssier, C., Campos, J., Jean-Pierre, H., Roger, F., Gay, B., Carlier, J. P., Jumas-Bilak, E. (2010). “Negativicoccus succinicivorans gen. nov., sp. nov., isolated from human clinical samples, emended description of the family Veillonellaceae and description of Negativicutes classis nov., Selenomonadales ord. nov. and Acidaminococcaceae fam. nov. in the bacterial phylum Firmicutes”. Int J Syst Evol Microbiol 60: 1271-9. doi:10.1099/ijs.0.013102-0. PMID 19667386.
- ^ Montgomery, L., Flesher, B., Stahl, D. (1988). “Transfer of Bacteroides succinogenes (Hungate) to Fibrobacter gen. nov. as Fibrobacter succinogenes comb. nov. and Description of Fibrobacter intestinalis sp. nov.”. Int J Syst Evol Microbiol 38: 430-435. エラー: 不正なDOI指定です. PMID 18048718.
- ^ Barns, S. M., Cainm E. C., Sommerville, L., Kuske, C. R. (2007). “Acidobacteria phylum sequences in uranium-contaminated subsurface sediments greatly expand the known diversity within the phylum”. Appl. Environ. Microbiol. 73 (9): 3113–6. doi:10.1128/AEM.02012-06. PMID 17337544.
- ^ Quaiser, A., Ochsenreiter, T., Lanz, C., et al (2003). “Acidobacteria form a coherent but highly diverse group within the bacterial domain: evidence from environmental genomics”. Mol. Microbiol. 50 (2): 563–75. doi:10.1046/j.1365-2958.2003.03707.x. PMID 14617179.
- ^ Eichorst, S. A., Breznak, J. A., Schmidt, T. M. (2007). “Isolation and characterization of soil bacteria that define Terriglobus gen. nov., in the phylum Acidobacteria”. Appl. Environ. Microbiol. 73 (8): 2708–17. doi:10.1128/AEM.02140-06. PMID 17293520.
- ^ Fukunaga, Y., Kurahashi, M., Yanagi, K., Yokota, A., Harayama, S. (2008). “Acanthopleuribacter pedis gen. nov., sp. nov., a marine bacterium isolated from a chiton, and description of Acanthopleuribacteraceae fam. nov., Acanthopleuribacterales ord. nov., Holophagaceae fam. nov., Holophagales ord. nov. and Holophagae classis nov. in the phylum ‘Acidobacteria’.”. Int J Syst Evol Microbiol 58: 2597-2601. エラー: 不正なDOI指定です. PMID 18984699.
- ^ Hou, S., Makarova, K. S., Saw, J. H., Senin, P., Ly, B. V., Zhou, Z., Ren, Y., Wang, J., Galperin, M. Y., Omelchenko, M. V., Wolf, Y. I., Yutin, N., Koonin, E. V., Stott, M. B., Mountain, B. W., Crowe, M. A., Smirnova, A. V., Dunfield, P. F., Feng, L., Wang, L., Alam, M. (2008). “Complete genome sequence of the extremely acidophilic methanotroph isolate V4, Methylacidiphilum infernorum, a representative of the bacterial phylum Verrucomicrobia”. Biology Direct 3. エラー: 不正なDOI指定です. PMID 18593465.
- ^ Saiki T, Kobayashi Y, Kawagoe K, Beppu T. (1985). “Dictyoglomus thermophilum gen. nov., sp. nov., a Chemoorganotrophic, Anaerobic, Thermophilic Bacterium”. Int J Syst Evol Microbiol 35: 253-259. エラー: 不正なDOI指定です.
- ^ 生命の黎明期に分岐した未知の菌株ジェマティモナス オーランティアカ(Gemmatimonas aurantiaca)の発見
- ^ Zhang, H., Sekiguchi, Y., Hanada, S., Hugenholtz, P., Kim, H., Kamagata, Y., Nakamura, K. (2003). “Gemmatimonas aurantiaca gen. nov., sp. nov., a Gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov.”. Int J Syst Evol Microbiol 53 (Pt 4): 1155-63. エラー: 不正なDOI指定です. PMID 12892144.
- ^ Cho, J.-C., Vergin, K. L., Morris, R. M. & Giovannoni, S. J. (2004). “Lentisphaera araneosa gen. nov., sp. nov, a transparent exopolymer producing marine bacterium, and the description of a novel bacterial phylum, Lentisphaerae”. Environmental Microbiology 6 (6): 611-21. エラー: 不正なDOI指定です. PMID 15142250.
- ^ Jumas-Bilak, E., Carlier, J. P., Jean-Pierre, H., Citron, D., Bernard, K., Damay, A., Gay, B., Teyssier, C., Campos, J., Marchandin, H. (2007). “Jonquetella anthropi gen. nov., sp. nov., the first member of the candidate phylum ‘Synergistetes’ isolated from man”. Int J Syst Evol Microbiol 56 (Pt 12): 2743-8. エラー: 不正なDOI指定です. PMID 18048718.
- ^ Jumas-Bilak, E., Roudière, L., Marchandin, H. (2009). “Description of ‘Synergistetes’ phyl. nov. and emended description of the phylum ‘Deferribacteres’ and of the family Syntrophomonadaceae, phylum ‘Firmicutes’”. Int J Syst Evol Microbiol 59 (Pt 5): 1028-35. エラー: 不正なDOI指定です. PMID 18048718.
- ^ Mori, K., Yamaguchi, K., Sakiyama, Y., Urabe, T., Suzuki, K. (2009). “Caldisericum exile gen. nov., sp. nov., an anaerobic, thermophilic, filamentous bacterium of a novel bacterial phylum, Caldiserica phyl. nov., originally called the candidate phylum OP5, and description of Caldisericaceae fam. nov., Caldisericales ord. nov. and Caldisericia classis nov.”. Applied and Environmental Microbiology 59 (Pt11): 2894-8. doi:10.1099/ijs.0.010033-0. PMID 19628600.
- ^ Geissinger O, Herlemann DP, Mörschel E, Maier UG, Brune A (2009). The ultramicrobacterium "Elusimicrobium minutum" gen. nov., sp. nov., the first cultivated representative of the termite group 1 phylum. 75. pp. 2831-40. doi:10.1128/AEM.02697-08. PMID 19270135.
- ^ Herlemann DP, Geissinger O, Ikeda-Ohtsubo W, Kunin V, Sun H, Lapidus A, Hugenholtz P, Brune A (2009). Genomic analysis of "Elusimicrobium minutum," the first cultivated representative of the phylum "Elusimicrobia" (formerly termite group 1). 75. pp. 2841-9. doi:10.1128/AEM.02698-08. PMID 19270133.
- ^ Hongoh Y, Sharma VK, Prakash T, Noda S, Taylor TD, Kudo T, Sakaki Y, Toyoda A, Hattori M, Ohkuma M (2008). Complete genome of the uncultured Termite Group 1 bacteria in a single host protist cell. 105. pp. 18391199. doi:10.1073/pnas.0801389105. PMID 19270133.
- ^ http://www.aist.go.jp/aist_j/press_release/pr2011/pr20110601/pr20110601.html 産業技術総合研究所はプレスリリース『新しい「門」アルマティモナデテス』]
- ^ Tamaki H, Tanaka Y, Matsuzawa H, Muramatsu M, Meng XY, Hanada S, Mori K, Kamagata Y. (2011). Armatimonas rosea gen. nov., sp. nov., of a novel bacterial phylum, Armatimonadetes phyl. nov., formally called the candidate phylum OP10.. 61. p. 1442-7.
[[Category:真正細菌]] [[en:List of Bacteria genera]] [[zh:細菌分類表]]

本記事は、古細菌の全分類群の一覧である。
古細菌は、他生物とは進化系統や細胞膜が大きく異なることから、3ドメイン説において生物の最上位のタクソンに位置付けられる。分離の困難さや進化速度の遅さなどの要因により、真正細菌(約10000)や真核生物(数100万)と比較して種の数は限られるものの、これまで高熱や強酸、高塩、嫌気的環境などの極限環境を中心に、約100属320種の古細菌が発見されてきた。
古細菌の学名は国際細菌命名規約に従い命名される。従って、厳密には細菌学名承認リストまたはIJSEMに記載された学名のみが有効である。分類は形状や生化学的性質よりも、DNAやRNAによる系統解析が重要視される傾向がある。
なお、一部の未記載種についても引用符をつけて掲載する。
概要(ドメインから綱まで)
[編集]- 真正細菌ドメイン/"Bacteria"
- 真核生物ドメイン/Eucarya (Eukaryota)
- 古細菌ドメイン/Archaea
- #ユリアーキオータ界/"Euryarchaeota"
- #ユリアーキオータ門/"Euryarchaeota"
- #メタノバクテリウム綱/Methanobacteria
- #メタノコックス綱/Methanococci
- #メタノミクロビウム綱/"Methanomicrobia"
- #ハロバクテリウム綱/Halobacteria
- #テルモプラズマ綱/Thermoplasmata
- #テルモコックス綱/Thermococci
- #アルカエオグロブス綱/Archaeoglobi
- #メタノピュルス綱/Methanopyri
- #ユリアーキオータ門/"Euryarchaeota"
- #クレンアーキオータ界/"Crenarchaeota"
- #クレンアーキオータ門/"Crenarchaeota"
- #テルモプロテウス綱/Thermoprotei
- #クレンアーキオータ門/"Crenarchaeota"
- 所属不明(クレンアーキオータ?)
- #タウムアーキオータ門/"Thaumarchaeota"
- #アイグアーキオータ門/"Aigarchaeota"
- #コルアーキオータ界/"Korarchaeota"
- #コルアーキオータ門/"Korarchaeota"
- 所属不明(ユリアーキオータ?)
- #ナノアーキオータ門/"Nanoarchaeota"
- #ユリアーキオータ界/"Euryarchaeota"
メタン菌。水素やギ酸を利用。動物の消化器官から水田、沼地、熱水噴出孔まで幅広く分布。細胞壁はシュードムレイン(グルム陽性)。
- メタノバクテリウム目/Methanobacteriales
- メタノバクテリウム科/Methanobacteriaceae
- MethanobacteriumAL
- Methanobacterium formicicum - M. aarhusense - M. alcaliphilum - M. beijingense - M. bryantii - M. congolense - M. espanolae - M. ivanovii - M. oryzae - M. palustre - M. subterraneum - M. thermaggregans - M. uliginosum - M. veterum
- Methanobrevibacter
- Methanobrevibacter ruminantium - M. acididurans - M. arboriphilus - M. curvatus - M. cuticularis - M. filiformis - M. gottschalkii - M. millerae - M. olleyae - M. oralis - M. smithii - M. thaueri - M. woesei - M. wolinii
- Methanosphaera
- Methanosphaera cuniculi - M. stadtmanae
- Methanothermobacter
- Methanothermobacter thermautotrophicus - M. defluvii - M. marburgensis - M. thermoflexus - M. thermophilus - M. wolfeii
- MethanobacteriumAL
- メタノテルムス科/Methanothermaceae
- Methanothermus
- Methanothermus fervidus - M. sociabilis
- Methanothermus
- メタノバクテリウム科/Methanobacteriaceae
メタン菌。水素やアミン類を利用。海洋や熱水鉱床、醗酵物など。細胞壁はS層(たんぱく質)。
- メタノコックス目/Methanococcales
- メタノコックス科/Methanococcaceae
- MethanococcusAL
- Methanococcus vannielii - M. aeolicus - M. maripaludis - M. voltae
- Methanothermococcus
- Methanothermococcus thermolithotrophicus - M. okinawensis
- MethanococcusAL
- メタノカルドコックス科/Methanocaldococcaceae
- Methanocaldococcus
- Methanocaldococcus jannaschii - M. fervens - M. indicus - M. infernus - M. vulcanius
- Methanotorris
- Methanotorris igneus - M. formicicus
- Methanocaldococcus
- メタノコックス科/Methanococcaceae

メタン菌。水田や動物の消化器官などに生息。比較的低温に適応。酢酸を利用できる種が多い。
- メタノミクロビウム目/Methanomicrobiales
- メタノミクロビウム科/Methanomicrobiaceae
- Methanomicrobium
- Methanomicrobium mobile
- Methanoculleus
- Methanoculleus bourgensis - M. chikugoensis - M. hydrogenitrophicus - M. marisnigri - M. palmolei - M. receptaculi - M. submarinus - M. thermophilus
- Methanofollis
- Methanofollis tationis - M. aquaemaris - M. ethanolicus - M. formosanus - M. liminatans
- Methanogenium
- Methanogenium cariaci - M. frigidum - M. marinum - M. organophilum
- Methanolacinia
- Methanolacinia paynteri
- Methanoplanus
- Methanoplanus limicola - M. endosymbiosus - M. petrolearius
- Methanosphaerula
- Methanosphaerula palustris
- Methanomicrobium
- メタノコルプスクルム科/Methanocorpusculaceae
- Methanocorpusculum
- Methanocorpusculum parvum - M. bavaricum - M. labreanum - M. sinense
- Methanocorpusculum
- メタノスピリルム科/Methanospirillaceae
- MethanospirillumAL
- Methanospirillum hungatei - M. lacunae
- MethanospirillumAL
- 未分類
- Methanocalculus
- Methanocalculus halotolerans - M. chunghsingensis - M. pumilus - M. taiwanensis
- Methanolinea
- Methanolinea tarda
- "Methanoregula"
- "Methanoregula boonei"
- Methanocalculus
- メタノミクロビウム科/Methanomicrobiaceae
- メタノサルキナ目/Methanosarcinales
- メタノサルキナ科/Methanosarcinaceae
- MethanosarcinaAL
- Methanosarcina barkeri - M. acetivorans - M. baltica - M. lacustris - M. mazei - M. semesiae - M. siciliae - M. thermophila - M. vacuolata
- Halomethanococcus
- Halomethanococcus doii
- Methanimicrococcus
- Methanimicrococcus blatticola
- Methanococcoides
- Methanococcoides methylutens - M. alaskense - M. burtonii
- Methanohalobium
- Methanohalobium evestigatum
- Methanohalophilus
- Methanohalophilus mahii - M. halophilus - M. portucalensis
- Methanolobus
- Methanolobus tindarius - M. bombayensis - M. oregonensis - M. profundi - M. taylorii - M. vulcani - M. zinderi
- Methanomethylovorans
- Methanomethylovorans hollandica - M. thermophila
- Methanosalsum
- Methanosalsum zhilinae
- MethanosarcinaAL
- メタノサエタ科/Methanosaetaceae
- Methanosaeta
- Methanosaeta concilii - M. harundinacea - M. thermophila
- Methanosaeta
- Methermicoccaceae
- Methermicoccus
- Methermicoccus shengliensis
- Methermicoccus
- メタノサルキナ科/Methanosarcinaceae
- メタノケッラ目/Methanocellales(ライスクルスターI)
- メタノケッラ科/Methanocellaceae
- Methanocella
- Methanocella paludicola - M. arvoryzae
- Methanocella
- メタノケッラ科/Methanocellaceae
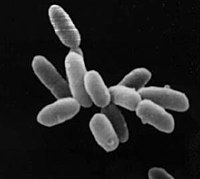
いわゆる高度好塩菌。塩湖・塩田などに分布。好気・光/化学合成従属栄養。
- ハロバクテリウム目/Halobacteriales
- ハロバクテリウム科/Halobacteriaceae
- HalobacteriumAL
- Halobacterium salinarum - H. jilantaiense - H. noricense - H. piscisalsi
- Haladaptatus
- Haloaptatus paucihalophilus - H. cibarius - H. litoreus
- Halalkalicoccus
- Halalkalicoccus tibetensis - H. jeotgali
- Halarchaeum
- H. acidiphilum
- Haloarcula
- Haloarcula vallismortis - H. amylolytica - H. argentinensis - H. hispanica - H. japonica - H. marismortui - H. quadrata
- Halobaculum
- Halobaculum gomorrense
- Halobiforma
- Halobiforma haloterrestris - H. lacisalsi - H. nitratireducens
- HalococcusAL
- Halococcus morrhuae - H. dombrowskii - H. hamelinensis - H. qingdaonensis - H. saccharolyticus - H. salifodinae - H. thailandensis
- Haloferax
- Haloferax volcanii - H. alexandrinus - H. denitrificans - H. elongans - H. gibbonsii - H. larsenii - H. lucentense - H. mediterranei - H. mucosum - H. prahovense - H. sulfurifontis
- Halogeometricum
- Halogeometricum borinquense - H. rufum
- Halogranum
- H. rubrum
- Halomicrobium
- Halomicrobium mukohataei - H. katesii
- Halonotius
- H. pteroides
- Halopelagius
- H. inordinatus
- Halopiger
- Halopiger xanaduensis - H aswanensis
- Haloplanus
- Haloplanus natans - H. vescus
- Haloquadratum
- Halorhabdus
- Halorhabdus utahensis - H. tiamatea
- Halorubrum
- Halorubrum saccharovorum - H. aidingense - H. alkaliphilum - H. arcis - H. californiense - H. chaoviator - H. cibi - H. coriense - H. distributum - H. ejinorense - H. ezzemoulense - H. kocurii - H. lacusprofundi - H. lipolyticum - H. litoreum - H. luteum - H. orientale - H. sodomense - H. tebenquichense - H. terrestre - H. tibetense - H. trapanicum - H. vacuolatum - H. xinjiangense
- Halosarcina
- Halosarcina pallida - H. limi
- Halosimplex
- Halosimplex carlsbadense
- Halostagnicola
- Halostagnicola larsenii - H. kamekurae
- Haloterrigena
- Haloterrigena turkmenica - H. daqingensis - H. hispanica - H. jeotgali - H. limicola - H. longa - H. saccharevitans - H. salina - H. thermotolerans
- Halovivax
- Halovivax asiaticus - H. ruber
- Natrialba
- Natrialba asiatica - H. aegyptia- H. chahannaoensis - H. hulunbeirensis - H. magadii - H. taiwanensis
- Natrinema
- Natrinema pellirubrum - H. altunense - H. ejinorense - H. gari - H. pallidum - H. versiforme
- Natronoarchaeum
- N. mannanilyticum
- Natronobacterium
- Natronobacterium gregoryi
- Natronococcus
- Natronococcus occultus - H. amylolyticus - H. jeotgali
- Natronolimnobius
- Natronolimnobius baerhuensis - H. innermongolicus
- Natronomonas
- Natronomonas pharaonis - N. moolapensis
- Natronorubrum
- Natronorubrum bangense - H. aibiense - N. sediminis - H. sulfidifaciens - H. tibetense
- "Haloalcalophilium"
- "H. atacamensis"
- HalobacteriumAL
- ハロバクテリウム科/Halobacteriaceae

好気性の好酸菌。細胞壁が無く、細胞融合性を持つことが特徴。硫黄孔、陸上の温泉などに分布。
- テルモプラズマ目/Thermoplasmales
- フェロプラズマ科/Ferroplasmaceae
- Acidiplasma
- Acidiplasma aeolicum - A. cupricumulans
- Ferroplasma
- Ferroplasma acidiphilum
- Acidiplasma
- ピクロフィルス科/Picrophilaceae
- Picrophilus
- Picrophilus oshimae - P. torridus
- Picrophilus
- テルモプラズマ科/Thermoplasmaceae
- 未分類
- フェロプラズマ科/Ferroplasmaceae
超好熱菌。何れも硫黄依存又は発酵性の嫌気従属栄養菌。海洋熱水噴出孔などに分布。
- テルモコックス目/Thermococcales
- テルモコックス科/Thermococcaceae
- Thermococcus
- Thermococcus celer - T. acidaminovorans - T. aegaeus - T. aggregans - T. alcaliphilus - T. atlanticus - T. barophilus - T. barossii - T. celericrescens - T. chitonophagus - T. coalescens - T. fumicolans - T. gammatolerans - T. gorgonarius - T. guaymasensis - T. hydrothermalis - T. kodakarensis - T. litoralis - T. pacificus - T. peptonophilus - T. profundus - T. sibiricus - T. siculi - T. stetteri - T. thioreducens - T. waiotapuensis - T. zilligii
- Palaeococcus
- Palaeococcus ferrophilus - P. helgesonii
- Pyrococcus
- Pyrococcus furiosus - P. glycovorans - P. horikoshii - P. woesei
- Thermococcus
- テルモコックス科/Thermococcaceae

超好熱菌。硫酸還元菌。微弱なメタン生成能有り。油田鉱床などに分布。
- アルカエオグロブス目/Archaeoglobales
- アルカエオグロブス科/Archaeoglobaceae
- Archaeoglobus
- Archaeoglobus fulgidus - A. lithotrophicus - A. profundus - A. sulfaticallidus - A. veneficus
- Ferroglobus
- Ferroglobus placidus
- Geoglobus
- Geoglobus ahangari - G. acetivorans
- Archaeoglobus
- アルカエオグロブス科/Archaeoglobaceae
メタノピュルス綱
[編集]超好熱性のメタン菌。水素を利用。海底熱水噴出孔。生育限界は120℃を超える。
- メタノピュルス目/Methanopyrales
- メタノピュルス科/Methanopyraceae
- Methanopyrus
- メタノピュルス科/Methanopyraceae
エオサイト界とも呼ばれる。殆どが超好熱菌または好熱好酸菌(スルフォロブス目)で、陸上温泉や海洋熱水系から分離される。原核生物の細胞分裂に関与するFtsZ分裂リング関連遺伝子の多くを持たず、一部を除き真核生物型のヒストンを持たない。なお伸長因子1αや大量データによる系統解析、rRNAのオペロン構造などから、真核生物がこの系統から派生した可能性が議論されている。後述のタウムアーキオータを含む可能性もある。
テルモプロテウス綱
[編集]
- テルモプロテウス目/Thermoproteales
- テルモプロテウス科/Thermoproteaceae
- Thermoproteus
- Thermoproteus tenax - T. neutrophilus - T. uzoniensis
- Caldivirga
- Caldivirga maquilingensis
- Pyrobaculum
- Pyrobaculum islandicum - P. aerophilum - P. arsenaticum - P. calidifontis - P. oguniense - P. organotrophum
- Thermocladium
- Thermocladium modestius
- Vulcanisaeta
- Vulcanisaeta distributa - V. souniana
- Thermoproteus
- テルモフィルム科/Thermofilaceae
- テルモプロテウス科/Thermoproteaceae
- アキディロブス目/Acidilobales
- アキディロブス科/Acidilobaceae
- Acidilobus
- Acidilobus aceticus - A. saccharovorans
- Acidilobus
- カルディスパエラ科/Caldisphaeraceae
- Caldisphaera
- Caldisphaera lagunensis
- Caldisphaera
- アキディロブス科/Acidilobaceae
- スルフォロブス目/Sulfolobales
- スルフォロブス科/Sulfolobaceae
- SulfolobusAL
- Sulfolobus acidocaldarius - S. metallicus - S. shibatae - S. solfataricus - S. tokodaii - S. yangmingensis
- Acidianus
- Acidianus infernus - A. ambivalens - A. brierleyi - A. sulfidivorans
- Metallosphaera
- Metallosphaera sedula - M. hakonensis - M. prunae
- Stygiolobus
- Stygiolobus azoricus
- Sulfurisphaera
- Sulfurisphaera ohwakuensis
- Sulfurococcus
- Sulfurococcus mirabilis - S. yellowstonensis
- SulfolobusAL
- スルフォロブス科/Sulfolobaceae
- デスルフロコックス目/Desulfurococcales
- デスルフロコックス科/Desulfurococcaceae
- Desulfurococcus
- Desulfurococcus mucosus - D. amylolyticus - D. fermentans - D. mobilis
- Aeropyrum
- Aeropyrum pernix - A. camini
- Ignicoccus
- Ignicoccus islandicus - I. hospitalis - I. pacificus
- Ignisphaera
- Ignisphaera aggregans
- Staphylothermus
- Staphylothermus marinus - Staphylothermus hellenicus
- Stetteria
- Stetteria hydrogenophila
- Sulfophobococcus
- Sulfophobococcus zilligii
- Thermodiscus
- Thermodiscus maritimus
- Thermosphaera
- Thermosphaera aggregans
- "Acidococcus"
- "Acidococcus sulfurreducens"
- "Fervidococcus"
- "Fervidococcus fontis"
- "Thermofermentum"
- "Thermofermentum shockii"
- Desulfurococcus
- ピュロディクティウム科/Pyrodictiaceae
- Pyrodictium
- Pyrodictium abyssi
- Hyperthermus
- Hyperthermus butylicus
- Pyrolobus
- "Geogemma"
- "Geogemma indica" - "G. pacifica" - "G. barossii"
- Pyrodictium
- フェルウィディコックス目/Fervidicoccales
- フェルウィディコックス科/Fervidicoccaceae
- フェルウィディコックス属/Fervidicoccus
- Fervidicoccus fontis
- フェルウィディコックス属/Fervidicoccus
- フェルウィディコックス科/Fervidicoccaceae
- 未分類
- "Caldococcus"
- "Caldococcus noboribetus"
- "Caldococcus"
- デスルフロコックス科/Desulfurococcaceae
"Cenarchaeum symbiosum"などのゲノム情報に基づいて2008年に提唱された門[2]。クレンアーキオータに含まれていた海洋性の古細菌(marine group 1 Crenarchaeaota)及び亜硝酸古細菌 (AOA:Ammonia-Oxidizing Archaea)がタウムアーキオータに分類された。系統的にはクレンアーキオータの深い部分から分岐したと見られ、低温への適応など他のクレンアーキオータとは異なった性質を持つ。培養が困難で純粋培養は"Nitrosopumilus maritimus"のみであり、2009年時点では記載種を含んでいない。
- ケナルカエウム目/Cenarchaeales
- "ケナルカエウム科"/"Cenarchaeaceae"
- "Cenarchaeum"(海綿に共生[3])
- "Cenarchaeum"(海綿に共生[3])
- "ケナルカエウム科"/"Cenarchaeaceae"
- "ニトロソプミルス目"/"Nitrosopumilales"
- "ニトロソプミルス科"/"Nitrosopumilaceae"(海洋性、水族館)
- "Nitrosopumilus"(水族館から発見。亜硝酸古細菌[4])
- "ニトロソプミルス科"/"Nitrosopumilaceae"(海洋性、水族館)
- "ニトロソカルドゥス目"/"Nitrosocaldales"
- "ニトロソカルドゥス科"/"Nitrosocaldaceae"
- "Nitrosocaldus"(好熱性亜硝酸古細菌[5])
- "Nitrosocaldus yellowstonii"
- "Nitrosocaldus"(好熱性亜硝酸古細菌[5])
- "ニトロソカルドゥス科"/"Nitrosocaldaceae"
- 所属不明
- "Nitrososphaera gargensis"

超好熱菌。イエローストーン国立公園の熱水底泥から検出された16S rRNA配列から前述の2界に属さない古細菌の存在が推測され提唱されたもの[6]。純粋培養はされていないが、2008年にゲノム自体は解読されている[7]。クレンアーキオータにやや近縁ともされる。
- "Korarchaeum cryptofilum"
メタゲノム解析により地下320mの金鉱から発見された好熱菌。ユビキチン-プロテアソームシステムに必要な遺伝子を持つ[8]
超好熱菌。Ignicoccusの表面に共生[9]しており、こちらも純粋培養はされていない。ユリアーキオータに含まれる可能性がある。2003年にゲノム解析自体は終了。ゲノムサイズは全古細菌中最小である(49万885塩基対)[10]。
関連項目
[編集]参考文献
[編集]- Boone, David R., Castenholz, Richard W., Garrity, George M. (2001). Bergey's Manual of Systematic Bacteriology, 2nd Edition, Volume One,The Archaea and the Deeply Branching and Phototrophic Bacteria. Springer-Verlag. ISBN 0387987711
- List of Prokaryotic names with Standing in Nomenclature
- Woese CR, Kandler O, Wheelis ML (1990). “Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya”. Proc Natl Acad Sci U S A 87 (12): 4576–9. PMID 2112744.
- ^ Gribaldo S, Brochier-Armanet C (2006). “The origin and evolution of Archaea: a state of the art”. Philos Trans R Soc Lond B Biol Sci 361 (1470): 1007–22. PMID 16754611.
- ^ Brochier-Armanet C, Boussau B, Gribaldo S, Forterre P. (2008). “Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota”. Nature Reviews Microbiology 6 (3): 245–52. PMID 18274537.
- ^ Preston CM, Wu KY, Molinski TF, DeLong EF (1996). “A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov”. Proc Natl Acad Sci U S A 93 (13): 6241–6. PMID 8692799.
- ^ Ingalls AE, Shah SR, Hansman RL, Aluwihare LI, Santos GM, Druffel ER, Pearson A (2006). “Quantifying archaeal community autotrophy in the mesopelagic ocean using natural radiocarbon”. Proc Natl Acad Sci U S A 103 (17): 6442–7. PMID 16614070.
- ^ de la Torre JR, Walker CB, Ingalls AE, Könneke M, Stahl DA. (2008). “Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol”. Environ Microbiol 103 (3): 810–8. PMID 18205821.
- ^ Barns SM, Delwiche CF, Palmer JD, Pace NR. (1996). “Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences”. Proc Natl Acad Sci U S A 93 (17): 9188–9193. PMID 8799176.
- ^ Elkins JG, Podar M, Graham DE, Makarova KS, Wolf Y, Randau L, Hedlund BP, Brochier-Armanet C, Kunin V, Anderson I, Lapidus A, Goltsman E, Barry K, Koonin EV, Hugenholtz P, Kyrpides N, Wanner G, Richardson P, Keller M, Stetter KO. (2008). “A korarchaeal genome reveals insights into the evolution of the Archaea”. Proc Natl Acad Sci U S A 105 (23): 8102–7. PMID 18535141.
- ^ Nunoura T, Takaki Y, Kakuta J, Nishi S, Sugahara J, Kazama H, Chee GJ, Hattori M, Kanai A, Atomi H, Takai K, Takami H. (2010). “Insights into the evolution of Archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group.”. Nucleic Acids Res. 39 (8): 3204-23. PMID 21169198.。系統的には、比較的[[タウムアーキオータに近いと考えられている。
- ^ Huber H, Hohn MJ, Rachel R, Fuchs T, Wimmer VC, Stetter KO (2002). “A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont”. Nature 417 (6884): 63–7. PMID 11986665.
- ^ Waters E, Hohn MJ, Ahel I, Graham DE, Adams MD, Barnstead M, Beeson KY, Bibbs L, Bolanos R, Keller M, Kretz K, Lin X, Mathur E, Ni J, Podar M, Richardson T, Sutton GG, Simon M, Soll D, Stetter KO, Short JM, Noordewier M (2003). “The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism”. Proc Natl Acad Sci U S A 100 (22): 12984–8. PMID 14566062.
[[Category:古細菌|ふんるい]] [[Category:分類学|こさいきん]]