Madarak
| Evolúciós időszak: Késő kréta–Jelen [1] | ||||||||||
 A madárvilág változatossága
| ||||||||||
| Rendszertani besorolás | ||||||||||
| ||||||||||
| Rendek | ||||||||||
| Hivatkozások | ||||||||||
A Wikimédia Commons tartalmaz Madarak témájú médiaállományokat és Madarak témájú kategóriát. |
A madarak (Aves) meszes héjú tojással szaporodó, melegvérű gerinces állatok. Mellső végtagjaik szárnyakká módosultak. Járólábaik vannak, melyeken megtalálható a jellegzetes csüd (lábtő és lábközépcsontok összenövése), amely elősegíti a landolást. Szegycsontjuk tarajos, melyhez erős mellizmok tapadnak, így segítve a repülést. A madárfajok nagy része ma is repülő életmódot folytat. Testüket tolltakaró borítja, csontjaik üregesek, légzsákokat tartalmaznak; ettől erősek, de könnyűek. Fogak helyett csőrük van. Méretük az 5 cm-es méhkolibritől a 2,75 m-es struccig terjed.
A legrégebbi ismert madárszerű faj az Archaeopteryx (ősgyíkmadár), amelynek csőre, karma, tolla, a lábán szarupikkelyek voltak (korábban ezt a fajt is az Avialae csoportba sorolták, ahogy a Theropodákat is, de azóta nem tekintik a madarak ősének). Őseik a Theropoda dinoszauruszok Maniraptora kládjába tartozó állatok voltak, ezáltal az utolsó fennmaradt dinoszauruszoknak tekinthetők. Legközelebbi élő rokonaik a krokodilok. A madarak mintegy 10 ezer fajjal a gerincesek második legnépesebb osztálya. Ennek több mint fele énekesmadár. Ebből Magyarországon közel 400 faj fordul elő.
A legtöbb faj szociálisan monogám, még akkor is, ha a párok nem mindig hűségesek. A párkapcsolat többnyire egy tenyészszezonra szól, néha több évig is eltart, de élethosszig ritkán. Néhány faj poligám, ezek lehetnek poligün fajok (egy hím, több nőstény), vagy poliandriát gyakorlók (egy nőstény, több hím). A nőstényeket tojóknak nevezik. Tojásait a legtöbb faj fészekbe rakja, de azok a fajok is saját testmelegükkel költik ki fiókáikat, amelyek nem építenek fészket. A legtöbb faj kikelés után még továbbgondozza fiókáit. A házi tyúk képes megtermékenyítetlen tojásokat is tojni.
Több madárfaj is fontos az ember számára; ezek egy része háziasított. Táplálékot (hús, tojás) és nyersanyagot (toll) biztosítanak. Egyes fajokat házikedvencként tartanak. Egyes fajok ürülékével trágyáznak (guanó). Az ember tevékenysége miatt a 17. századig több száz, a 17. századtól 100-120 faj halt ki, és további 1200 faj veszélyeztetett. Megfigyelésük az ökoturizmus fontos része.
Törzsfejlődés
[szerkesztés]Rokonság
[szerkesztés]
A madarakat először Francis Willughby és John Ray rendszerezte 1676-ban kiadott Ornithologiae művükben.[3] Carl Linnaeus ezt felhasználva alkotta meg saját rendszerét 1758-ban.[4] Az ő nyomán a madarakat osztályként tartják nyilván, filogenetikailag a Theropoda kládba sorolhatók.[5]
A madarak és testvércsoportjuk, a krokodilok az Archosauria klád ma is élő tagjai. Az 1990-es évek végéig az Archaeopteryx lithographica összes utódját a madarak közé sorolták.[6] Jacques Gauthier inkább csak a modern madarakat tekintette valódi madárnak, és a csak fosszíliákból ismert fajokat a madarakkal együtt a bővebb Avialae csoportba sorolta.[7] Ezzel elkerülte az Archaeopteryx körüli bizonytalanságot. A 21. században ez az osztályozás terjedt el.
Gauthier[8] négyféle meghatározást adott a madarakra. Ezzel azonban az a probléma, hogy nem ugyanazok a fajok tartoznak oda. Továbbá azt javasolta, hogy a madarakat a ma élő összes madár közös ősétől számítsák. Az általa meghatározott csoportok:
- tollas archosaurusok (Avifilopluma)
- röpképes archosaurusok (Avialae)
- azok az archosaurusok, amelyek közelebb állnak a ma élő madarakhoz, mint a krokodilokhoz (Avemetatarsalia [=Paraves])
- a ma élő madarak legközelebbi közös ősének leszármazottai (Neornithes)
| ||||||||||||||||||
| A madarak filogenetikai rokonsága a ma is élő hüllőkkel |
A negyedik definíció szerint az Archaeopteryx nem tartozik a madarak közé. Gauthier javaslatát a legtöbb paleontológus és a madarak evolúciójával foglalkozó szakértők is elfogadták, habár pontos meghatározás nincs. A kutatók gyakran az Avialae tudományos névvel illetett csoportot nevezik madaraknak.[9]
Az Avialae a legtöbb kutató szerint ág-alapú klád, habár a meghatározások eltérnek egymástól. Sok szerző értelmezésében azok a theropodák, amelyek közelebb állnak a madarakhoz, mint a Deinonychushoz.[10][11] Néha testi jellemzők alapján különítik el a kládot. Jacques Gauthier 2001-ben új definíciót adott: tollas lények, amik legalább siklórepülésre (voltak) képesek, köztük a tőlük leszármazó madarakkal.[8][12]
Kapcsolat a dinoszauruszokkal
[szerkesztés]
| ||||||||||||||||||||||||||||||
| Kladogram Cau et al., 2015 filogenetikai eredményei alapján[13] |
A madarak az Archosauriák fejlődési irányához tartozó hüllőmedencéjű dinoszauruszok (Saurischia) egy csoportjának (Theropoda) ágából alakultak ki. A valódi madarak a kréta korban, 100 millió évvel ezelőtt jelentek meg.[14] DNS-elemzések szerint nagyobb mértékben a többi dinoszaurusz kihalása után terjedtek el a déli kontinensről világszerte. Fajszámuk különösen a globális jégkorszakok alatt gyarapodott.[15] Primitív madárszerű dinoszauruszok a középső juráig visszakövethetők. A tollas dinoszauruszokból alakult ki a legkorábbi ismert madárszerű lény, az Archaeopteryx. Ezek még nem repültek jól, és őseik testfelépítéséből még megőrizték a fogakkal teli állkapcsot, és a hosszú csontos farkat.[16][17]
A fosszíliák tanúsága és a biológiai bizonyítékok miatt a legtöbb tudós a madarakat specializálódott theropodáknak tartja.[18] Speciálisabban, a Maniraptora klád része; ide tartoznak többek között a dromaeoszauridák és az oviraptoridák is.[19] Ahogy egyre több, madárhoz hasonló theropoda került elő, elmosódtak a határok a madarak és rokonaik között. Északkelet-Kínában Liaoning tartományban a sok apró tollas theropoda került elő, ami tovább növelte a kétséget.[20][21][22]

A paleontológia 2010-es évekbeli állása szerint a repülő theropodák, azaz madárszárnyúak (Avialae) a deinonychoszauruszok legközelebbi rokona, és magukba foglalják a dromaeoszauridákat és a troodontidákat is.[23] Ezek együtt alkotják a Paraves csoportját. Több emblematikus tag, mint a Microraptor, olyan jellemzőkkel bír, amelyek lehetővé tették számukra legalább a siklórepülést. A legtöbb emblemetaikus tag kis termetű volt. Ez felveti azt a lehetőséget, hogy a paravianok közös őse fán élt, és képes volt a siklórepülésre.[24][25] Az Archaeopteryxtől és a többi theropodáktól eltérően az első madárszárnyúak (Avialae) nem ragadozók vagy dögevők, hanem mindenevők voltak.[26]
Az Archaeopteryx a 19. században az egyik megkerült hiányzó láncszemnek és az evolúció egyik bizonyítékának számított. Ez volt az első lelet, amin látszódtak még a hüllők jellegzetességei: fogak, karmos mellső ujjak, hosszú, hüllőszerű farok, és a modern madarakhoz hasonló tollas szárnyak. Ma már nem tekintik a madarak közös ősének, de hozzá közel állónak igen.[27]
Az első madárszerű állatok
[szerkesztés]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kladogram Cau et al., 2015 filogenetikai eredményei alapján[13] |
A legkorábbi avialan fosszíliákat Kínában, a Tiaojishan Formációban fedezték fel, és a jura szakaszára datálták, 160 millió évvel ezelőttre.[9] Az ismert akkori avialan fajok: Anchiornis huxleyi, Xiaotingia zhengi, és Aurornis xui. A jól ismert Archaeopteryx valamivel későbbi, 155 millió éves, és Németországban találták. Az ősi, dinoszauruszokra utaló jegyek mellett még további jellegzetességeik voltak, mint a második lábujj karma, ami sosem érintette a földet; és a hosszú tollak, amikkel a levegőben manővereztek.[28]
Az Avialae fajokból sok különböző forma alakult ki a kréta korban.[29] Több csoport megtartotta a korai jegyeket, míg más csoportok elvesztették ezek egyikét-másikát. Így például a Pygostylia csoportban megrövidült a farok, és létrejött a pygostyl csont.[29] A késői krétában 95 millió évvel ezelőtt a modern madarak őseinek megjavult a szaglása.[30]
A korai madárősök sokfélesége
[szerkesztés]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Egyszerűsített mezozoós madár filogenia Wang et al. szerint, 2015-ös filogenetikai elemzés alapján[31] |
A rövid farkúakból kialakultak az enantiornithesek, melyek vállcsontjainak szerkezete fordított volt a modern madarakéhoz képest. Ők voltak az első változatos csoport a rövid farkúak közül. Sokféle ökológiai nichét foglaltak el, a homokban gázolóktól és halevőktől a falakó magevőkig. A kréta korban ők voltak a legelterjedtebb madárszerűek, azonban a kor végén kihaltak.[29]
A másik ág, az Euornithes vízi vagy félig vízi életmódot folytattak, halakkal és más kisebb vízi élőlényekkel táplálkoztak; gázló, úszó és búvár fajaik voltak. Ide tartoztak a sirályszerű Ichthyornis,[32] és a röpképtelenné vált Hesperornithiformes, amelyek tengerekben halásztak.[29] Az enantiornithesek ezzel szemben a szárazföldön terjedtek el.[33] A modern madarak irányába mutat a csőr kifejlődése és a fogak elvesztése, habár voltak csoportjaik, amelyek megőriztek néhány hátsó fogat.[34] Emellett mellcsontjukon taraj alakult ki. Farkcsontjuk mozgékonnyá vált, amivel a farok is szabadabban mozgott,[35] és lehetővé tette a repülés közbeni manőverezést.[28]
Modern madarak
[szerkesztés]
| |||||||||||||||||||||||||||
| A modern madarak főbb csoportjai Sibley-Ahlquist taxonómiája szerint |
A modern madarakat és őseiket az Aves, más néven Neornithes néven foglalják össze, aminek két ága van: (1) Futómadár-szabásúak (Palaeognathae), ahova az olyan röpképtelen madarak tartoznak, mint a struccalakúak (Struthioniformes) rendje, és az olyan rosszul repülők, mint a tinamualakúak (Tinamiformes) rendje; és (2) a tarajos szegycsontúak (Neognathae), ahová a többi madarat sorolják.[5] Ezeket többnyire nagyrendnek vagy alosztálynak tekintik,[36] de Livezey és Zusi cohorsként (raj) tartja számon.[5] A rendszertani nézőponttól függően a modern madarak ismert fajainak száma 9800[37] és 10 050 között változik.[38]
A récefélék családjába tartozó késő krétában élt Vegavis faj leletei alapján a madarak már a kréta kor végén több ágra váltak szét.[39] Alaktani elemzések szerint a modern madarak valamivel a legkorábbi fosszíliák előtt, a kréta kor közepén jelentek meg.[16]
Az ágak közül először a Galloanserae vált le, ami a lúdalakúak és a tyúkalakúak öregrendje. Valószínűleg tőlük származik a valódi madarak első fosszíliája is 85 millió évvel ezelőttről, az Austinornis lentus fajtól.[40] Az újabb szétválások időpontjai vitatottak; a kutatók csak abban értenek egyet, hogy az első Galloanserae fajok a késő krétabeli dinoszauruszok kortársai voltak. Abban viszont nincs egyetértés, hogy a többi faj mikor és milyen úton terjedt el.[41] A bizonyítékok ellentmondásosak; a molekuláris elemzések szerint még a kréta kor végén, míg a leletek szerint csak a kainozoikum alatt; ez vitathatóvá teszi az összes eredményt.[41][42] Mindenesetre az újabb eredmények azt mutatják, hogy a kréta kor végi kihalást csak kevés faj élte túl, és ezekből származik a mai madarak egy népes csoportja.[43]
Elterjedésük
[szerkesztés]
A madarak jelen vannak a hét kontinens legtöbb szárazföldi élőhelyén, egészen a hóhojsza költőtelepeiig, amelyek az Antarktiszon 440 kilométerre vannak a tengertől.[45] A fajok a trópusi területeken a legváltozatosabbak. Korábban úgy gondolták, hogy ennek oka az erősebb fajképződés, azonban azóta kiderült, hogy a fajképződés inkább a magasabb szélességeken gyorsabb, mivel ott nagyobb a fajkihalás is.[45] A szárazföldek mellett a vizeken is élnek, egyes családok még az óceánokban is előfordulnak. Némely tengerlakó faj csak költeni megy a partra,[46] és néhány pingvinfaj akár 300 méterre is lemerül az óceánban.[47]
Sok madárfaj az ember segítségével terjedt el. Ez lehetett akaratlagos, vadnak (fácán)[48] vagy véletlen, elszabadult díszállatként, mint a barátpapagáj Észak-Amerika egyes városaiban.[49] Néhány faj hasznot húzott az ember tevékenységéből, így a pásztorgém,[50] a pásztorkarakara,[51] vagy a rózsás kakadu, és elszaporodott a mezőgazdasági területeken[52] vagy a városokban.
Testfelépítés
[szerkesztés]A madarak, bár igen változatos állatcsoport, testfelépítés szempontjából viszonylag egységesek. Más gerincesekhez képest több szokatlan jellegzetességgel bírnak, amelyek elsősorban a repülőképesség kialakulásához kapcsolódnak.
Idegrendszer, érzékelés
[szerkesztés]A madarak központi idegrendszere viszonylag fejlett.[53] Az agy mozgással foglalkozó kérgében a repülést irányító rész a legnagyobb. A kisagy vezérli a pontos mozgásokat, míg a nagyagy a komplex viselkedésmintákat, például a tájékozódást, az udvarlást és a fészeképítést. Az idegrendszer egyes részeinek megnevezése új terminológiát igényelt, mivel a madáragy másként épül fel, mint az emlősöké.[54] Az 1990-es évek óta végzett vizsgálatok szerint a madarak értelmesebbek, mint ahogy azt korábban gondolták.
A madáragyra jellemző a sok apró idegsejt; térfogatához és tömegéhez képest sok sejtből áll. Az intelligensebb fajok (énekesmadarak, papagájok) agya tömegarányosan sokkal több sejtet tartalmaz, mint az emlősöké, a főemlősöket is beleértve. Testtömeghez képest az idegsejtek száma magasabb, mint a legtöbb emlősben, a főemlősöket kivéve. Például a sárgafejű királyka testtömege kilencede egy egérének, mégis kétszer annyi idegsejtje van. Az idegsejtek a nagyagyban, különösen a nagyagy kérgében helyezkednek el sűrűn, ami énekesmadaraknál és papagájoknál három-négyszer annyi idegsejtet jelent, mint a főemlősökben. A madárfajok között óriási a különbség az agy viszonylagos méretét illetően. A bankivatyúk testtömege ötvenszerese a széncinegének, mégis ugyanannyi neuronja van. De még a tyúkalakúak és a struccok esetén is akkora az idegsejtek sűrűsége, mint a főemlősöknél. A papagájok és az énekesmadarak nagy agya az utódgondozásnak köszönhető. A fejlettebb utódgondozás, mint feladat egy összetett viselkedésminta lehetővé teszi, hogy a fiókák agya is nagyra nőjön.[55]
Látás
[szerkesztés]A madarak látása általánosságban jól fejlett. A vízimadarak szemlencséje képes egyaránt alkalmazkodni a víz alatti és a víz feletti viszonyokhoz, és éles látást biztosítani.[53] Egyes fajoknak két sárgafoltja van. Négyféle csaptípusuk van, a vörös, zöld, kék mellett ultraibolya fényt észlelő csapjaik is vannak (tetrakromázia).[56] Az ultraibolya fény látása segíti őket a táplálékszerzésben és a párkeresésben is. A szem mellett agyukban is vannak fényérzékeny sejtek, amelyek észlelik a nappalok hosszának változását, és vezérlik az évszaknak megfelelő viselkedést.[57]
Egyes madárfajok, például a kék cinke tollazata ultraibolya mintákat is mutat. Ez segíti a nemek megkülönböztetését. A hím udvarlás közben ezeket mutatja különböző pózok felvételével és a tollak felborzolásával. A sólymok a rágcsálók által hagyott vizeletjelzések segítségével találják meg zsákmányukat. A galambok és néhány más faj kivételével a madarak nem a szemhéjukkal, hanem pislogóhártyával pislognak, ami vízszintesen mozog. Vízimadarak ezt kontaktlencseként használják a vízben. A madarak szemében a fésűszerv gondoskodik a tápanyagellátásról és szabályozza az üvegtest kémhatását. A legtöbb madár nem tudja mozgatni a szemét; ha máshova akar nézni, akkor a fejét kell elmozdítania. Ez alól csak néhány kivétel van, például a kormorán. Azok a madarak, amelyeknek oldalt van a szemük, fejük elfordítása nélkül is látnak hátrafelé; azok a madarak, amelyeknek elöl van a szemük, távolságot tudnak becsülni, mint például a baglyok.
Hallás
[szerkesztés]A madaraknak nincs külső füle, csak néhány fajnál alkotnak fülszerű mintázatot a tollak, például egyes baglyoknál. A középfülben csak egy hallócsont van. A belső fülben csiga is van, viszonylag rövid és nincs feltekeredve. A baglyok füle aszimmetrikus, így pontosabban meg tudják határozni a leendő zsákmány helyét. A baglyok hallását arcuk alakja és tollazatának elrendeződése is segíti.[58] Ezzel szemben a legtöbb madárnak el kell mozdítania a fejét, hogy meghatározza a hangforrás irányát. Körülbelül ugyanazokat a hangokat hallják, mint az ember, a 100 Hz alatti hangok kivételével. A hallás időbeli felbontása kiváló, de érzékenyebb az erős zajokra. Egyes fajok akár négy hangot is képesek egyszerre kiénekelni.
Egyensúlyozás
[szerkesztés]Nemcsak fülükben van egyensúlyozó szervük, hanem a medencében is, ami a testhelyzetet elemzi, és segíti az egyensúlyozást például ágon üléskor. Ha ez a szerv tönkremegy, akkor a madár nem képes reagálni az ág helyzetének megváltozására.
Szaglás és ízlelés
[szerkesztés]A legtöbb madár szaglása kifejezetten gyenge, ami megközelítően az emberének felel meg. Ez alól kivételek a kivifélék,[59] az újvilági keselyűk,[60] és a viharmadár-alakúak, akik szaglással keresik táplálékukat.[61] Orrlyukaik a csőr tövén nyílnak.
Az ízlelőbimbók nem a nyelven, hanem a nyelv tövénél és a garatban helyezkednek el. Számuk alacsony az emlősökéhez képest (kacsa 200, ember 9000). A táplálékszerzésben alárendelt szerep jut az ízlelésnek, szerepe mégsem elhanyagolható.
Tapintás
[szerkesztés]A legtöbb madárnak a nyelve és a csőre a legérzékenyebb a tapintási ingerekre. Táplálékkereséskor a látást, táplálékfelvételben az ízlelést egészíti ki. Fára mászáskor a lábujjaik által észlelt ingerek alapján tájékozódnak. A lilealakúak és más madarak tapintással ismerik fel a táplálékukat, amikor az iszapban kotorásznak.
Mágneses érzék
[szerkesztés]Egyes fajok, különösen a vándormadarak észlelik a Föld mágneses terét, az erővonalak irányát. Ennek érzékszerve a szemben[62] (és/vagy) a csőrben helyezkedik el. Először Wolfgang Wiltschko mutatta ki vörösbegyen a Zoologischen Institut in Frankfurt am Mainben 1967-ben.
A szemben levő mágneses érzékszerv a gyökpárképződés elvén működik, ugyanis a szembe érkező fény hatására molekulák gyökökké bomlanak. Erre a folyamatra hathat a Föld mágneses tere. A csőrben mágneses részecskék alkotják az iránytűt; elmozdulásukat a környező idegszövet észleli. A technikai eszközökkel szemben ez az iránytű inkább az inklinációt méri.
Csontváz
[szerkesztés]
A madaraknak két koponyatípusát különböztetjük meg: a futómadarakra és tinamukra jellemző paleognath és a repülő madarakra, valamint a pingvinekre jellemző, fejlettebb felépítésű neognath koponyát. Kifejlett madarakon a koponyacsontok összenőttek.[53] Emellett a madarakra általánosan jellemző a fogatlan csőr és a nagy méretű szemüreg. A két szemüreget csontos sövény választja el.
A madarak csontjai, szemben az emlősök tömör csontjaival, üregesek, ezzel csökkentve az állat súlyát. A csontok nagyon könnyűek, üregeik a lélegzőrendszerhez kapcsolódnak.[63]
A madarak számos (11-24) nyakcsigolyája igen lazán, heterocoelikus módon kapcsolódik egymáshoz, így a madárnyak rendkívül mozgékony. A hát mozgása már korlátozott, a többi szakasz pedig mozdíthatatlanul összenőtt, és az úgynevezett álkeresztcsontot vagy ágyékkeresztcsontot hozták létre.[53][64]
A bordák laposak, és a futómadarak kivételével az összes madárfaj szegycsontján jellegzetes tarajt (crista) találunk, amely tapadási felületet biztosít a repülőizmoknak.[65] (A futómadarak közül egyedül a tinamualakúaknál található tarajos szegycsont, de ezek is röpképtelenek, vagy igen rosszul repülnek).
A mellső végtagban (szárny) elkülöníthető a felkarcsont, az orsócsont és a singcsont, valamint két kéztőcsont, de a többi kéztőcsont a kézközépcsontokkal összenőve hozza létre a carpometacarpust.
A hátsó végtagban a combcsontot, a tibiotarsust (a lábszárcsont és néhány lábtőcsont összenövéséből), a vékony szárkapocscsontot, a csüdcsontot és a lábujjperceket különíthetjük el. A legtöbb madárfajnak 4 lábujja van, de a futómadarak esetében ez redukálódhat 3 vagy 2 lábujjra.
Izomzat
[szerkesztés]
A madarak szárnyát hatalmas méretű mellizmok (musculus pectoralis, musculus supracoracoideus) mozgatják, amelyek a szegycsont taraján tapadnak meg. A tarajról indulva az egyik izom (a m. pectoralis) a felkarcsont alsó felületéhez, a másik (m. cupracoracoideus) a vállízületet megkerülve a felkarcsont felső felületéhez tapad. Vagyis a szárny lecsapását és felemelését végző izmok ugyanott erednek.
A madarak lábának különleges izma a musculus pectinus, amely a térdízülethez tapad, és inakkal az egyes lábujjakhoz csatlakozik. Ha a madár behajlítja a térdét, a m. pectinus megfeszül, és a madár lábujjai összezáródnak, ezzel biztosítva a kapaszkodást.
Kültakaró
[szerkesztés]
A madarak testét tollak borítják, amelyeket evezőtollakra, fedőtollakra és pehelytollakra különítünk el. Ez ma már csak a madarakra jellemző; a földtörténeti múltban a nem a valódi madarakhoz tartozó dinoszauruszok között is voltak tollasak. Ez biztosítja a hőszigetelést, és segít a repülésben is. A tollak még fontos szerepet játszanak a párválasztás során, illetve az álcázásban.[53] Több különböző tolltípus létezik a különféle célokra. A legtöbb madáron pásztákban nőnek, csak a pingvinek, a tüskésszárnyúmadár-félék és a ratites fajokon oszlanak el egyenletesen.[66] A pászták eloszlása jellemző a különféle rendszertani egységekre, így segíthetnek a meghatározásban. A tollazat egy fajon belül is különböző lehet, függ a nemtől,[67] a kortól és a társadalmi helyzettől.[68]

Rendszeresen vedlenek. Az évente kétszer vedlő fajok költési időn kívül nyugalmi tollruhát, udvarlási és költési időben nászruhát viselnek. A legtöbb faj évente vedlik; a nagy ragadozók még ritkábban, több évente. A vedlési mintázat fajonként változó. Az énekesmadaraknál a szárnytollak vedlése szimmetrikusan halad belülről kifelé. A következő toll akkor hullik ki, miután az előző már kinőtt. A fedőtollak az alattuk levő tollakkal együtt kerülnek sorra. A belső primary az első; miután az első öt primary lecserélődött, azután a terciary tollak következnek. Utánuk a secondary tollak jönnek.[69] Néhány faj, például a kacsák egyszerre vesztik el szárnytollaikat, így egy időre röpképtelenekké válnak.[70] A faroktollak vedlési sorrendje a legtöbb fajon hasonló, belülről halad kifelé,[69] de például a fácánoknál kívülről kezdődik.[71] Harkályokon némileg módosult, a kifelé haladó fázis kihagyja a középső tollakat, azok a végén cserélődnek, így a harkály közben is támaszkodhat rá.[69][72] Énekesmadarakon a primary tollak kifelé, a secondary tollak kívülről befelé, és a farok belülről kifelé cserélődik.[73] Költés előtt a legtöbb faj nőstényei megkopaszodnak a hasukon. A bőr gazdagon erezett, ez segít a költésben.[74]
A madarak többnyire minden nap tollászkodnak, ébren töltött idejük 9%-át fordítják erre a tevékenységre.[75] Eltávolítják az idegen testeket. A madarak bőre mirigyekben szegény. Legfontosabb mirigyük a farcsíkmirigy (Glandula uropygii), amelynek zsírban gazdag viaszos váladéka vízhatlanná teszi a tollazatát, és akadályozza a baktériumok növekedését és szaporodását.[76] Egyes madaraknál különösen fejlett, más fajoknál hiányozhat. Tollászkodás végén ezzel a váladékkal keni be magát a madár. Ezt kiegészítheti hangyasavas kezeléssel, amit a hangyáktól szerezhet meg.[77]
A madarak lábán a pikkelyek ugyanúgy keratinból vannak, mint a csőr, a tollak és a karmok. Többnyire az ujjakon és a metatarsuson láthatók, de egyes madarakon (jégmadár, fakopáncsok) bokáig terjednek. Úgy gondolják, hogy homológok a hüllők és az emlősök pikkelyeivel.[78]
Emésztőrendszer
[szerkesztés]A madarak nyelőcsövének jellegzetes tágulata a begy, amelynek két típusa létezik: a két oldalra irányuló begyzsákokból álló valódi begy és a nyelőcső egyszerű tágulata, az álbegy. Előbbi a galambokra, énekesmadarakra és tyúkfélékre, utóbbi a ragadozó madarakra és a récefélékre jellemző. Több ragadozó madár köpetet képez a zsákmány maradékából.[79]
A madarak gyomra két részre tagolódik, a mirigyes gyomorra és az izmos zúzógyomorra vagy zúzára, amivel a táplálékot megőrlik, mivel nincs foguk, mert az akadályozná őket a repülésben. A madaraknál ezért előfordul, hogy homokot vagy apró kavicsokat nyelnek le, hogy azok segítsék őket a táplálék összezúzásában. A legtöbb faj a repülő életmódhoz való alkalmazkodás miatt gyorsan emészt.[80] Egyes vándormadarak fehérjében tárolnak tartalékokat a vándorlás idejére.[81]
Tápcsatornájuk rendre kloákában végződik, amely a tápcsatorna, az ivarszervek és a húgycső közös kivezetőnyílása.
Légzés
[szerkesztés]A szárazföldi állatok közül a madaraknál alakult ki a legbonyolultabb és leghatékonyabb légzőszervrendszer.[53] Kettős légzéssel lélegeznek, melynek lényege, hogy a többi állattal ellentétben nemcsak be-, hanem kilégzéskor is történik gázcsere.[82] Ezt elsősorban a tüdő speciális felépítése teszi lehetővé. Belégzéskor a levegő 75%-a a tüdőbe, a maradék 25% a légzsákokba kerül.
További érdekesség, hogy a madarak hangképző szerve nem a gégében, hanem a főhörgők elágazásánál található, ezért két helyen is képződik hang. Az alsó gégefők izmos kamrák, amelyek többszörös membránt tartalmaznak.[83] Egyes fajokban a légcső a testhez képest lényegesen hosszabb, ami felerősíti a madár hangját.[84] A madár ily módon képes egyszerre énekelni és levegőt venni, illetve egyszerre több hang kiadására is képes lehet. (A fülemülék például egyszerre négy különböző hang kiadására is képesek.)
A tüdő felépítése
[szerkesztés]
A madarak tüdővel lélegeznek, melynek felépítése erősen eltér az emlősök tüdejétől. Míg az emlősök tüdejében a levegő vakon végződő léghólyagokba (alveolusokba) fut, addig a madarakéban úgynevezett léghajszálcsövek találhatók, melyeken a levegő folyamatosan keresztüláramolhat.
Szerkezet alapján kétféle tüdőt különíthetünk el: a primitívebb paleopulmót, és a fejlettebb neopulmót. A paleopulmóban a levegő kilégzéskor és belégzéskor is ugyanabba irányba áramlik, a neopulmóban ellentétesen.
A tüdőhöz 5 pár légzsák csatlakozik, amelyek hatékonyabbá teszik a ki- és belégzést, valamint könnyebbé teszik a madár testét. A madarak tüdejének térfogata állandó, a légcserét a légzsákok nyomásváltozása működteti.
A kettős légzés
[szerkesztés]A légcső (trachea) közvetlenül kapcsolódik a két hátulsó pár légzsákhoz valamint a paleo- és neopulmóhoz is. Belégzéskor a friss levegő közvetlenül átáramlik mind a paleo-, mind a neopulmón, így belégzéskor gázcsere történik. Ugyanekkor a hátulsó légzsákba is levegő kerül: egyrészt friss levegő közvetlenül a tracheából; másrészt a neopulmón átáramló, és már oxigénben szegényebb levegő is ide jut. Így a belégzés végére a légzsákban oxigénben viszonylag gazdag levegő lesz. Kilégzéskor a hátulsó légzsákokból a levegő egyrészt a légcsövön keresztül kiáramlik, másrészt ismét átáramlik a neopulmón így kilégzéskor is történik gázcsere. A légzsákból közvetlenül, és a neopulmón keresztül is kerül levegő a paleopulmóba, ezzel is növelve a gázcsere intenzitását.[82]
Keringés
[szerkesztés]A madarak az emlősökhöz hasonlóan állandó testhőmérsékletű, úgynevezett meleg vérű állatok. Szívük két-két, szilárd válaszfal által elkülönített kamrából és pitvarból áll, és rostos zsák veszi körül. A rostos zsákban a szív folyadékban úszik. A vér visszaáramlását a szívben billentyűk gátolják. A szívverés ütemét a jobb pitvar falában található szinuszcsomó adja meg. A depolarizációs jelet kalciumionok okozzák. A szív izomzata ívekbe rendeződik, amelyek rétegeket alkotnak, a belső, a középső és a külső rétegeket.[85] A pitvarok fala vékonyabb, mint a kamráké, mert a kamráknak erőteljesen kell összehúzódniuk az oxigéndús vér pumpálásához. Az azonos testtömegű emlősökhöz képest a madarak szíve nagy; így több vért tud egy időegység alatt pumpálni, hogy fedezze a repülés miatt megnőtt oxigén- és energiaszükségletet.[86]
Két vérkörük van, a jobb szívfél a kisvérköri, a bal szívfél a nagyvérköri keringést tartja fenn. A kis vérkörben a jobb kamrából a szén-dioxidos vér a tüdőartérián keresztül a tüdőbe áramlik, majd onnan oxigéndús vér áramlik vissza a bal pitvarba. A bal kamrából oxigéndús vér áramlik a test szövetei felé, onnan szén-dioxidban dús vér a jobb pitvarba. A madarakban nagyon hatékony a gázcsere; a tüdőben tízszer akkora felület áll ehhez rendelkezésre, mint az azonos méretű emlősöknek, és a tüdőben több vér van a kapillárisokban hosszegységenként.[86] Az artériákat a szívből kiindulva vastag elasztikus izmok védik a szívverés okozta nyomáskülönbségtől. A szívtől távolodva ahogy vékonyodnak, egyre merevebbek lesznek, amelyek oxigént és tápanyagokat szállítanak az egész testnek.[87] Az egyre kisebb ágakra oszló artériák megnövelik a felszínt, és a vér folyása lelassul. A vér befolyik a legkisebb kapillárisokba, itt lezajlik a gázcsere. A vér annyira lelassul, hogy a gázcsere a lehető leghatékonyabb legyen. Az oxigéndús vér a bal pitvarba, majd a bal kamrába folyik. Az oxigénszegénnyé vált vér kis vénákba, majd vénákba gyűlik, és visszajut a szívbe, ahol is a jobb pitvarba, majd jobb kamrába ömlik.[87]
Kiválasztás
[szerkesztés]A madarak kiválasztószervei a hüllőkhöz hasonlóan működnek. Veséjük a nitrogéntartalmú salakanyagot húgysavként választja ki.[88][89][90] Néhány madár, például a kolibrik képesek egy másik anyagcsereúton ammónia formájában is megszabadulni a fölös nitrogéntől.[91] Az emlősöktől eltérően kreatinin helyett kreatint választanak ki a kloákába, ami a végbél utolsó szakasza.[53] Nincs húgyhólyagjuk; a vizelet a végbélbe folyik (a strucc kivételével), majd onnan félig folyékony állapotban az ürülékkel távozik.[92][93] A végbélnek ez a szakasza a kloáka, nőstényekben a tojások is ezen haladnak át.
Szaporodó szervek
[szerkesztés]A madarak két neműek, vannak hímek és nőstények. Nemüket a szexkromoszómák határozzák meg az ivart meghatározó ZW rendszer szerint. Eszerint a nőstényeknek ZW, a hímeknek ZZ kromoszómáik vannak.[53] Tehát a nem megtermékenyüléskor eldől, a hőmérsékletnek erre nincs hatása. Azonban előfordulhat, hogy az egyik nem érzékenyebb a melegre, ami nemtől függő halandóságot okoz.[94]
A madarak belső megtermékenyítésűek. A legtöbb faj a kloákák összeérintésével párzik. Néhány fajnál a hímeknek van péniszük, ilyenek a lapos szegycsontúak a kivik kivételével, a lúdalakúak a tüskésszárnyúmadár-félék kivételével, és a tyúkalakúak. A Neoaves többi csoportjánál nincs pénisz.[95][96] A pénisz méretét a spermaverseny erősségével indokolják.[97] Párzáson kívül a pénisz a kloákában pihen.[98]
Ez az egyetlen gerinces csoport, ahol az elevenszülés semmilyen formában nem jelent meg. Minden madárfaj meszes héjú tojásokat rak.
Védekezés
[szerkesztés]A viharmadár-alakúak néhány faja képes gyomrából olajat felöklendezni támadóira.[99] Új-Guineában a pitohuis fajok közül egyesek tollai és bőre idegmérget tartalmaz.[100]
Néhány fajnak éles sarkantyúja van a szárnyán; ezek a tüskésszárnyúmadár-félék, fácánfarkú levéljáró, tüskésszárnyú lúd, zuhatagi réce és kilenc bíbicfaj. Néhány más faj csontos bütyköt visel vagy viselt a szárnyában, ezek: a gőzhajórécék (Tachyeres), lúdformák, Rodriguez-szigeti galamb (kihalt), Chionis, Crax és Burhinus fajok. A kihalt jamaikai íbisz szárnya hosszú volt, és kézfej is volt rajta.[forrás?] Ezeket a fegyvereket használhatják a ragadozók vagy akár saját fajtársaik ellen is, amivel súlyos sérüléseket vagy akár halált is okozhatnak.[101]
Életmód
[szerkesztés]A legtöbb faj nappal aktív, éjjel és szürkületkor csak a baglyok és a lappantyúk aktívak. A tengerparti madarak nem az éjjel-nappali ciklushoz, hanem az árapályhoz igazodnak, és éjjel és nappal is járnak táplálkozni, ha a vízállás megfelelő.[102]
Repülés
[szerkesztés]
A madarak alkalmazkodtak a repüléshez. A legtöbb madár tud repülni, habár vannak röpképtelen fajok. A mellső végtag szárnnyá alakult.[53] A szárnyak alakja és mérete határozza meg, hogyan repül a madár. Általában erős szárnyverdeséses szakaszok váltakoznak siklórepüléssel. Az erős repülőizmok eredése a mellcsonton kiemelkedő tarajon található. A testtömeg 15%-át teszik ki.[53] Tollaik a bőr származékai, a szárny- és faroktollak a repülésre és a kormányzásra szolgálnak. Szivacsos szerkezetűek a csontjaik, ezért könnyebbek. A repülés nagy izommunkát igényel, ezért van szükségük fejlett légzésre és keringési rendszerre. Mindezek megkülönböztetik őket a többi gerincestől.
A repülést felhasználják táplálékszerzéshez, fészeképítő anyagok kereséséhez, ragadozók elkerülésére és a ragadozók elől való menekülésre. Voltak szigetek, ahová a madarak eljutottak, de az emlős ragadozók nem.[103] Ezeken a szigeteken a madarak az emlősök szerepét betöltve és a szűkös erőforrások miatt röpképteleneké váltak. Hatvan jelenleg is élő faj és több kihalt faj is röpképtelen (volt).[104] Egyes fajok ugyan nem repülnek, de repülő mozdulatkat végeznek úszás, búvárkodás közben: pingvinek, alkák. Ezeknek a repülő fajokhoz hasonló izomzatuk van. Puffin fajok és vízirigók úgy úsznak, ahogy repülnek.[105] Néhány faj az energiában szegény táplálék feldolgozásához alkalmazkodott. A futómadarak a gyors futásért váltak röpképtelenekké.
A repülés fizikai alapjai
[szerkesztés]
A madarak repülését a szárny felépítése teszi lehetővé. A madarak szárnya repülés közben úgy tereli a levegőt, hogy a szárny felett gyorsabban áramoljon, mint alatta. Így a felül áramló levegő nyomása kisebb lesz. A madár az ebből adódó aerodinamikai felhajtóerőt használja fel a repüléshez.
A madárszárny (és a legtöbb repülő állat szárnyának) különlegessége, hogy lecsapáskor a szárnyprofil az állat sebességétől függően változtatható, így képes a rá ható felhajtóerőt változtatni. A madarak repülése rengeteg kutatót és mérnököt foglalkoztat. Ez a mechanizmus ugyanis sokkal hatékonyabb, mint a repülőgépekben alkalmazott, mivel ezek szárnyprofilját csak nagyon kis mértékben lehet változtatni.
A madarak között egyedülálló a kolibrifélék repülési technikája. Szárnyaikkal előre-hátra csapkodva nyolcast írnak le a levegőben, így képesek egy helyben lebegni. (Erre egyetlen más madár sem képes.) Ehhez rendkívül gyorsan kell verdesniük: a legkisebb kolibrik másodpercenként akár hetven szárnycsapásra is képesek.
Intelligencia
[szerkesztés]Egyes madarakat, mint a varjúféléket és a papagájokat a legértelmesebb állatok között tartják számon. Több faj is készít és használ eszközöket, továbbá tudást örökít át, amit kultúrának is lehet tekinteni. A vándormadarak minden évben nagy távolságokat tesznek meg. Társas állatok, hanggal és látható jelzésekkel kommunikálnak, és sok közös tevékenységben vesznek részt, mint telepeken költés, közös vadászat, tollászkodás és ragadozók elkergetése.
Táplálkozás
[szerkesztés]
A különböző madárfajok táplálkozása változatos. Fogyasztanak nektárt, gyümölcsöt, növényeket, magokat, de dögöt és más állatokat is, gerincteleneket és gerinceseket egyaránt, köztük más madárfajokat.[53] Mivel nincsenek fogaik, emésztőrendszerük alkalmazkodott a rágás nélkül megevett táplálék gyors emésztéséhez.[106]
Vannak generalisták, akik többféle módszerrel többféle táplálékot fogyasztanak, míg a specialisták csak egyféle módszert használnak, és erősen válogatósak.[53] Sok faj gerincteleneket, gyümölcsöket és magokat szedeget, de vannak fajok, amelyek lesből támadva kapják el apró zsákmányukat. A kártevőket elfogyasztó madarakat szívesen látják a gazdaságokban.[107] A nektárevők (kolibrifélék, nektármadárfélék, lórifélék) koevolválódtak táplálékadó növényeikkel, és ecsetszerű nyelvük van.[108] Kivifélék és gázlómadarak hosszú csőrükkel keresik gerinctelen zsákmányukat; a gázlómadarak fajonként változó hosszúságú csőre és módszerei különböző ökológiai fülkékre utalnak.[53][109] A búvármadarak, bukórécék, pingvinek és alkák víz alatt üldözik zsákmányukat, míg a szulafélék, jégmadárfélék, csérfélék légi vadászok, amelyek a levegőből csapnak bele a vízbe. A flamingófélék, három prionfaj, és néhány kacsa a vizet szűri.[110][111] A libák és a réceformák főként legelnek.
Egyes madarak, mint a fregattmadárfélék, a sirályfélék[112] és a halfarkasfélék[113] kleptoparaziták, azaz más madaraktól lopnak élelmet. Viszont nem ez a fő táplálékforrásuk; a halászattal, vadászattal szerzett élelmüket egészítik ki. Egy kutatás a nagy fregattmadarak álarcos szuláktól való lopásait vizsgálta. A cikk szerint átlagban csak 5%-ot lopnak, és ebből táplálékuk legfeljebb 40%-a származik.[114] Egyes fajok dögöt esznek; vannak, amelyek emellett vadásznak is, mint a sirályok, varjúfélék és más ragadozók, és vannak, amelyek nem, ezek a keselyűk.[115]
Ivás
[szerkesztés]A madarak vízigénye viszonylag kicsi, mivel nincsenek izzadtságmirigyeik és kiválasztásuk is hatékonyabb, mint az emlősöké.[116] Egyes sivatagi madarak képesek csak a táplálékból fedezni folyadékszükségletüket. Emellett másként is adaptálódtak ehhez a biotóphoz, például túlélik, ha testhőmérsékletük megemelkedik, és összegyűjtik a harmatot.[117] A tengeri madarak tengervizet isznak, a sót pedig a fejükön elhelyezkedő mirigyek választják ki.[118]
A legtöbb madár úgy iszik, hogy először a csőrébe veszi a vizet, majd felemeli a fejét, hogy a víz lefolyjon a nyelőcsövén. A galambfélék, díszpintyfélék, egérmadárfélék, guvatfürjfélék és túzokalakúak tudnak a fejük felemelése nélkül is inni.[119] Néhány sivatagi madár függ a vizesgödröktől, és a pusztaityúk-félék seregestül járnak vízért. A hímek hastollaikba itatva visznek vizet családjuknak.[120] Vannak fajok, amelyek begyükben szállítják a vizet, és a táplálékkal együtt öklendezik fel. A galambok, flamingók, pingvinek begytejet választanak ki a fiókák táplálására.[121]
Tollászkodás
[szerkesztés]A tollászkodás fontos a madár egészsége szempontjából. Mivel a tollazat életfontosságú, rendben kell tartani. A tollakat gombák, tollatkák és madártetvek támadják.[122] A tollazat átkenése mellett a madarak szívesen fürödnek vízben vagy porban. Egyes fajok belemerülnek a vízbe, de vannak, amelyek inkább belerepülnek. Az erdei madarak gyakran a leveleken összegyűlt esővízben vagy harmatban mártóznak. Száraz területeken homokfürdőt vesznek. A hangyafürdőzés azt jelenti, hogy a madár hagyja, hogy hangyák árasszák el tollruháját. A hangyasav tovább tisztítja a tollakat és a bőrt. Egyes fajok kitárt szárnyakkal napfürdőznek.[123][124]
Vándorlás
[szerkesztés]
Sok faj kihasználja a különböző éghajlati öveket, és vándorol. A vándormadarak közé tartoznak szárazföldi, vízi és parti fajok, amelyek mérsékelt vagy hideg égövben szaporodnak, és költési időn kívül a trópusokra vagy a másik féltekére repülnek. Indulás előtt feltöltik raktáraikat, testzsírjuk megnövekszik, és egyes szerveik összemennek,[81][125] mivel a hosszú repülőutak nagyon energiaigényesek, különösen, ha tengereken vagy sivatagokon repülnek át, és nincs lehetőség táplálkozni. A szárazföldi vándormadarak többnyire körülbelül 2500, míg a parti madarak 4000 km-t tudnak megtenni.[126] A rekorder a kis goda, ami éjjel-nappal repülve 10 200 km-re jut el.[127] Tengeri vándormadarak még hosszabb utakra indulnak, a leghosszabb utat a Chilében és Új-Zélandon költő szürke vészmadár teszi meg, és Japánban, Alaszkában és Kaliforniában tölti a nyarat. Évente összesen 64 ezer kilométert utazik.[128] Más tengeri madarak inkább rögzített útvonal nélkül nagy távolságokra kóborolnak. A Déli-óceánon fészkelő albatroszok gyakran egészen az északi sarkvidékig repülnek.[129]
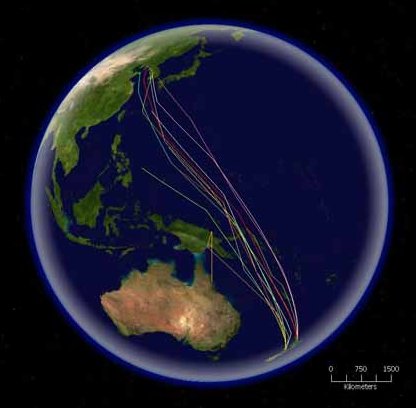
A többi faj is kóborol, azaz rögzített útvonal nélkül rövidebb távolságokat tesz meg, hogy elkerülje a rossz időjárást és táplálékot szerezzen. Ilyenek a pintyek, amelyek költőhelyükhöz sem ragaszkodnak; évről évre új helyet keresnek maguknak, így ha az egyik évben sokan vannak egy helyen, akkor a következőben lehet, hogy eltűnnek onnan. Előfordul, hogy ugyanannak a fajnak az északibb területein költő példányai oda húzódnak, ahol fajtársaik egész évben tartózkodnak.[130] Mások csak részben vándorolnak, általában nőstények és alávetett hímek.[131] Egyes régiókban sok faj viselkedik így; Ausztráliában a nem énekesmadarak körében 44%, míg énekeseknél 32%. Magassági vándorlásra hegyekben költő fajok indulnak, a rossz idő közeledtével lejjebb vándorolnak. Ezt többnyire a lehűlés váltja ki, és a hegyek lakói jobb táplálékellátottságú helyeket keresnek maguknak.[132] Néhány faj nomád életmódú, nem tart territóriumot, hanem mindig oda vonul, ahol megfelelőek a körülmények. A papagájok rövidebb utakat tesznek meg.[133]
Már régóta ismert, hogy a madarak visszatalálnak fészkelőhelyükre. Az 1950-es években Bostonban útjára bocsátottak egy atlanti vészmadarat, ami 13 nap alatt 5150 km távolságról visszatért kolóniájába a walesi Skomernál.[134] Vándorlásuk alatt a madarak többféleképpen is tájékozódnak. Nappal a Napot, éjjel a Holdat és a csillagokat használják iránytűnek. A Nap helyzetének változását belső órájuk segítségével kompenzálják.[53] A csillagok közül a poláris csillagképek helyzetéhez igazodnak.[135] Emellett egyes fajok a Föld mágnesességét is kihasználják.[136]
Kommunikáció
[szerkesztés]
A madarak látható és hallható jelekkel kommunikálnak. A jelzések egy része a fajtársakhoz, másik része más fajok tagjaihoz szól.
Testtartásukkal és tollazatuk felmeresztésével kifejeznek dominanciaigényt,[137] vagy fenyegetőznek, eljátszva a nagy ragadozót, mint például a guvatgém a ragadozókkal szemben, hogy megvédje fiókáit.[138] A tollazat formája, színe és mintázata a faj, a nem vagy az egyed azonosítását is lehetővé teszi. A látható kommunikáció magában foglalja a rituálékat is, amelyeknek eredetileg nem jelzésértékük volt, hanem gyakorlati eredményük. A rituálék jelezhetnek agressziót, behódolást, vagy a másik párként való elfogadását.[53] A legbonyolultabb rituálék az udvarlást kísérik; a táncok gyakran sok különböző mozdulatból állnak.[139] A hímek szaporodási sikere ezektől a táncoktól függ.[140]
Probléma esetén lásd:Médiafájlok kezelése.
A madarak hangképző szerve az alsó gégefő, ami a hörgők kettéágazásánál található. A hangzó kommunikáció hívásokból és énekekből áll, amelyek nagyon összetettek lehetnek. A madarászoknak le kell lassítaniuk a felvételeket, hogy értelmezni tudják a hallottakat. Egyes fajok külön tudják mozgatni a két gégefőt, így duettet tudnak énekelni önmagukkal.[83]
A hívójelek sokfélék. Hívják társaikat,[53] felmérik a közelben tartózkodókat,[141] párzásra hívnak, védik területüket,[53] azonosítják egymást;[142] ragadozókra figyelmeztetnek, néha a veszély típusát is megjelölve.[143] Néhány madár a saját hangját mechanikai hangokkal egészíti ki. A Coenocorypha fajai levegőt vezetnek át tollaikon,[144] a fakopáncsok fákon kopogtatva jelzik területüket,[80] és a pálmakakaduk eszközökkel dobolnak.[145]
Társas kapcsolatok
[szerkesztés]
Néhány faj területhez kötve, családban él, míg mások nagy rajokat alkotnak. A rajképzés előnye a nagy szám által adott védelem, és a hatékonyabb táplálékszerzés.[53] Az egy faj által alkotott nagy rajok biztonságot adnak, de növelik a versengést a táplálékért.[147] A ragadozók elleni védelem különösen fontos a zárt területeken, mint az erdők, ahol a lombok eltakarhatják a ragadozókat, de több szem többet lát, így hamarabb riadót fújhatnak. Ez nagy, sok fajt magukban foglaló csapatok kialakulásához vezetett, ahol a különböző fajok nem versengenek egymással a táplálékért, és az egyes fajok létszáma kicsi. Ugyanis, ha túl sokan lennének egy fajból, akkor az alacsony rangú tagok bántalmazása nagyban csökkentené a táplálékszerzés hatékonyságát.[148]
A madarak néha más, nem madár fajokkal is társulnak. A tengeri madarak gyakran vadásznak együtt delfinekkel és tonhalakkal, így a zsákmányhalaknak egyik irányból sincs menekvés.[149] A szarvascsőrűmadár-félék a közönséges törpemongúzzal társulnak, és együtt keresnek táplálékot, és kölcsönösen figyelmeztetik egymást a veszélyre.[150]
Pihenés
[szerkesztés]
Gyors anyagcseréjük miatt a madarak az aktív napszakukban is alszanak időnként. Alvás közben néha rövid időre kinyílik a szemük, hogy szemrevételezzék a környezetet, nincs-e veszély.[151] Úgy tudják, hogy a sarlósfecskefélék repülés közben is tudnak aludni, és radarmegfigyelések szerint a magasságcsökkenést a széllel szembe fordulva ellensúlyozzák.[152] Egyes alvástípusok alkalmasak lehetnek arra, hogy a madarak repülés közben is pihenhessenek.[153] Egyes fajoknál kimutatták, hogy agyféltekéik külön is tudnak aludni. Ezt a képességüket a rajban elfoglalt helyükhöz képest használhatják ki, mivel így az éber féltekéhez tartozó szem a kívülről érkező veszélyeket figyelheti. Tengeri emlősöknél is ismert hasonló adaptáció.[154] A madarak gyakran összegyűlnek éjszakára, aminek céljai a hatékony hőszabályozás és védekezés.[155] A gyülekezőhelyet is ennek megfelelően választják. Izzadságmirigyek híján különféleképpen hűtik magukat, például árnyékba húzódnak, vízbe állnak, kitárják szárnyukat, vagy levizelik magukat.[156]
Sok madár alvás közben nyakát a hátára hajtja, és csőrét a háttollak közé fúrja, míg más fajok a melltollaik közé dugják csőrüket. Sok madár fél lábon alszik, míg mások mindkét lábukat a tollaik közé rejtik, különösen hideg időben. A Loriculus nem papagájai fejjel lefelé lógnak.[157] Az énekesmadarak lába reflexesen zárul a gally körül alvás közben. Néhány nagyobb madárfaj, mint a fürj és a fácán éjszakára felgallyaz, vagyis fára települ alváshoz. Néhány kolibrifaj anyagcseréje jelentősen lelassul éjszakára.[158] Ez több száz faj esetén megtörténik, így a kuvikfecskealakúak, az Artamus fajok és a lappantyúfélék esetén is. A téli estifecske hibernálódik.[159]
Szaporodás
[szerkesztés]
A fajok 95%-a szociálisan monogám. Ez azt jelenti, hogy egy hím és egy nőstény párba áll, a legtöbb fajnál egy tenyészszezonra, néhány fajnál évekre vagy egy egész életre.[161] Ez lehetővé teszi, hogy a szülők közösen neveljék fiókáikat, mivel egy szülő csak nagy nehezen, vagy egyáltalán nem járna sikerrel.[162] Azonban a legtöbb fajnál a félrelépés sem ritka.[163] Ez tipikus, ha a nőstény domináns hímmel lép félre, vagy a réceféléknél a hím erőszakot követ el a nősténnyel.[164] A nőstények tartalékolni tudják a spermát, egyes fajok akár 100 napig is,[165] így több hím spermáját is összegyűjtheti, így bebiztosíthatja magát arra az esetre, ha a párjának nem működne a spermája.[166] Ezeknél a fajoknál a párzási időszakban a hímek kísérgetik párjukat, hogy nehogy megcsalja őket, és hogy valóban a saját fiókáikat neveljék fel.[167]
Vannak más szaporodási rendszerek is, előfordul promiszkuitás és a poligámia több változata is (egy hím, több nőstény; több hím, egy nőstény; több hím, több nőstény). A poligám rendszerek akkor alakulnak ki, ha egy szülő, többnyire a nőstény egyedül fel tudja nevelni a fiókákat.[53] Néhány fajnál több rendszer is előfordul, a körülményektől függően. Homoszexuális párokat is megfigyeltek hímeknél és nőstényeknél is, udvarlással, párba állással, párzással, félrelépéssel és közös utódneveléssel.[168]
A párba állást általában udvarlás előzi meg. Tipikusan a hím udvarol a nősténynek.[169] A legtöbb egyszerű, az udvarló énekel; más rendszerekben kifinomult táncot ad elő, repül, fészket vagy lugast épít, néha testvéreivel közösen, lekben.[170] Általában a nőstény választ párt, de ez alól is van kivétel: a víztaposó, ahol a nemek szerepe fordított. A hímek választanak párt a színpompás nőstények közül,[171] és a hímek nevelik fel a fiókákat. Egymás etetése az udvarlás alatt nem jellemző, általában párzás után fordul elő.[80]
Fészekrakás és költés
[szerkesztés]Sok faj területfoglalással védi a család táplálékbázisát. Amely fajok nem foglalnak területet, azok telepesen fészkelnek, és a sokaság nyújtotta védelmet élvezik. Ők a fészekhelyet védik, mivel a telep belseje védettebb, mint a külső része. Egyes telepeken több faj költ, itt a küzdelem független a fajtól.[172]

Többnyire az üregekben költő fajok fehér tojást, a szabadon álló fészekben költők terepszínű tojást raknak; ez alól azonban sok a kivétel. A lappantyúfélék halvány héjú tojásukat tollakkal álcázzák. Azok a fajok, amelyeknek költésparazitáik vannak, többféle színű tojást raknak, hogy észrevehessék a költésparazita tojását. Minden új szín előnyben van. A költésparaziták azzal alkalmazkodnak, hogy olyan fészket keresnek, ahol a tojások az övéihez hasonlóak.[173]


A legtöbb madár fészekbe rakja tojásait. A fészek alakja sokféle lehet, lehet csésze, dóm, lapos, emeletes, üreg vagy halom.[174] Néhány fajnak nincs szüksége jól kiépített fészekre; az albatroszok csak kimélyítenek egy gödröt, az a fészek. A legtöbb madár elrejti, de a telepesen költők inkább nyitott fészket építenek, mivel védettebbek. Egyes fajok olyan növényeket válogatnak össze, amelyek védenek az élősködőktől, hogy a fiókák egészségesek legyenek,[175] és hőszigetelésként gyakran építenek be tollakat.[174] Néhány faj egyáltalán nem épít fészket; a lumma a földre rakja a tojásait, a császárpingvin a lábai között tartja. A fészek hiánya gyakori azoknak a fajoknak a körében, amelyek talajra rakják tojásaikat, és fiókáik fészekhagyók.
A fészekhagyó madarak fiókái képesek követni anyjukat kikelésük után és önállóan táplálkoznak, de például repülni még nem tudnak. Általában talajszinten fészkelnek. A lúdalakúakra és a tyúkalakúakra jellemző.
A fészeklakó madarak általában csupaszon (csekély tollazattal), fejletlenül (csukott szemmel) kelnek ki, a szülők még sokáig gondozzák őket. Mivel a fiókák még nem önállóak és nem tudnak repülni, sőt, eleinte még járni sem, nagyrészt fákon fészkelnek, hogy az utódok védve legyenek a ragadozóktól. Kevés tojást raknak, mert a szülők nem tudnak sok fiókát gondozni.

A legtöbb faj az utolsó tojás lerakása után kezd kotlani.[53] A költés optimalizálja a hőmérsékletet a fiókák fejlődése számára. Monogám fajok megosztják a költés idején a teendőket, habár vannak fajok, ahol csak az anya kotlik, és az apa hordja a táplálékot. Poligám fajok esetén mindent egy szülő intéz. A hőt a has vagy a mell csupasz bőre közli. A költés energiaigényes folyamat; az albatroszok akár napi 83 grammot is veszítenek súlyukból a költés alatt.[176] Az ásótyúkfélék nem maguk kotlanak, hanem a Nap, a vulkánok, vagy bomló növényzet hőjét használják fel.[177] A költési idő 10 naptól (apró fajok, énekesmadarak, harkályok, kakukk) 80 napig (nagy ragadozó madarak, albatroszok és kiwik) terjed.[53]
Nagyok a különbségek a különböző, akár közeli rokon fajok között. Néhány faj fő jellemzőit az alábbi, rendezhető táblázat foglalja össze:[178][179]
| Faj | Felnőttkori testtömeg
(gramm) |
Költési idő
(nap) |
Költések száma
(évente) |
Fészekalj mérete |
|---|---|---|---|---|
| Tüzestorkú kolibri (Archilochus colubris) | 3 | 13 | 2,0 | 2 |
| Házi veréb (Passer domesticus) | 25 | 11 | 4,5 | 5 |
| Kaliforniai földikakukk (Geococcyx californianus) | 376 | 20 | 1,5 | 4 |
| Pulykakeselyű (Cathartes aura) | 2200 | 39 | 1,0 | 2 |
| Laysan-albatrosz (Diomedea immutabilis) | 3150 | 64 | 1,0 | 1 |
| Magellán-pingvin (Spheniscus magellanicus) | 4000 | 40 | 1,0 | 1 |
| Szirti sas (Aquila chrysaetos) | 4800 | 40 | 1,0 | 2 |
| Vadpulyka (Meleagris gallopavo) | 6050 | 28 | 1,0 | 11 |
Utódnevelés
[szerkesztés]Az utódnevelés hossza és természete erősen fajfüggő. Néhány faj, például az ásótyúkfélék már kikelésétől kezdve képes az önálló életre, és szülői segítség nélkül ássa ki magát a költődombból.[180] A másik szélsőséget a tengeri madarak képviselik, amelyeknél a szülői gondoskodás sokáig elhúzódik. A nagy fregattmadár fiókái hat hónap alatt tollasodnak ki, és hagyják el a fészket, de további tizennégy hónapig etetik őket.[181] A fiókákra vigyázó szülő segíti a hőgazdálkodást, és véd a ragadozókkal szemben.[182]

A nevelést végezheti egy vagy mindkét szülő. Néhány fajnál a már felnőtt előző (vagy még idősebb) fészekalj is bekapcsolódik testvéreik nevelésébe.[183] Megfigyelhető a Corvida ág tagjainál,[184] de megfigyelhető az álcsuszkánál és a vörös kányánál is. A madaraknál gyakori az apai gondoskodás a fiókákról.[53] Az apa részt vesz a terület és a fészek védelmében, a költésben és a fiókák etetésében. Néha a különböző feladatok megoszlanak az apa és az anya között.[185]
A kitollasodás és a fészek elhagyásának ideje változó. A Synthliboramphus fajok fiókái a kikelésüket követő éjszaka elhagyják a fészket, és a tengeren nevelkednek, távol a szárazföldi ragadozóktól.[186] A fészekhagyó fajok fiókái szüleik vezetésével korán elhagyják a fészket. A legtöbb fajnál a fészek elhagyása és a repülés között nem telik el sok idő, de ez erősen változó. Miután az albatroszok fiókái elhagyták a fészket, nem kapnak további segítséget, de sok más fajé igen.[187] A fiatalok követik szüleiket első vándorútjukon.[188]
Költésparazitizmus
[szerkesztés]
A költésparazitizmus azt jelenti, hogy a parazita az utódját becsempészi egy másik faj fészkébe, és azzal nevelteti fel. Ez a madarak körében a leggyakoribb.[189] A gazdafaj többnyire elfogadja sajátnak a parazita tojását, kikölti és felneveli a saját utódai kárára. Lehet feltételes vagy feltétlen. Ha feltétlen, akkor az anya az összes utódját más fajjal nevelteti fel, míg a másik esetben van saját fészke, de néhány tojást más fajokhoz csempész be, ha módja van rá.[190] Száznál több madárfajról tudják, hogy feltétlen költésparazita: az ismert kakukkokon kívül mézkalauzfélék, csirögefélék, récefélék közül a kakukkréce tartoznak ide. Néhány faj korábban kel ki, mint a gazdamadár fiókái, és kidobálja a tojásokat és a fiókákat, ha mégis megelőzték a kikeléssel. Más fajok utódai vagy kiéheztetik a többi fiókát, vagy velük együtt nevelődnek fel.[191]
Szexuális szelekció
[szerkesztés]
A madarak körében többféle párválasztási módszer alakult ki. A pávaformák farka a Fisher-féle megszaladási jelenség legismertebb példája. Az általános nemi kétalakúság, mint a méret és a színezet különbsége az energiaszükséglet iránti igény különbségét is okozzák. A nagyobb mértékű különbség különösen költséges jellemzők kialakulását jelenti; ezek versenyhelyzetet jeleznek.[192][192] A madarak körében többféle szexuális szelekció ismert; az egyik rendszerben a nőstények választanak a hímek közül; a másikban az egyik nem tagjai megküzdenek egymással a párosodás jogáért. A megszaladási jelenségek eredményeként az óriási szárnyak és farkak könnyen észrevehetővé teszik a madarat, és akadályozzák is a mozgást. Ez a konfliktus biztosítja, hogy a jelek igaziak; ezekkel a díszekkel csak a jó életképességű hímek képesek életben maradni.[193]
Beltenyészet és elkerülése
[szerkesztés]A beltenyészet közeli rokonok utódnemzését jelenti. Többnyire előnytelen, mert a közeli rokonok sok génnek ugyanazt az allélját hordozzák, így a recesszív allélok gyakrabban találnak egymásra, ami rontja a túlélés esélyeit.[194] Ugyanis ezek gyakrabban okoznak betegséget, mint a domináns allélok. Mivel egy-egy faj minden tagja többé-kevésbé rokon, azért nem rokonnal nem állhatnak párba, de ha a faj nem veszélyeztetett, akkor lehet távoli rokont találni.[195][196]
A beltenyészet a zebrapintynél korai halált okozhat.[197] A távoli rokon párokhoz képest a testvér-testvér pároknál szignifikánsan rosszabb volt a kelési arány.
A Geospiza scandens földi kaktuszpinty esetén a hatást környezeti tényezők is befolyásolják, például az elérhető táplálék mennyisége és minősége.[198]
A bíborsapkás tündérmadár esetén a közeli rokonok párzásából megtermékenyült tojások esetén több mint 30%-kal csökkent a kelési arány. Az ilyen kapcsolatokban a félrelépés is gyakoribb; az ilyen párok költéseinek 43%-ában találtak félrelépésből született fiókát. Ezzel csökkentik a beltenyészet káros hatásait.[199]
A széncinege esetén is rosszabbak a beltenyészetből származó utódok túlélési esélyei. A természetben a cinegék ezt azzal kerülik el, hogy születési helyüktől távol telepednek meg, így csökken annak az esélye, hogy közeli rokonukkal állnak párba.[200]
A tarka rigótimália képes felismerni közeli rokonait, és szintén eltávolodik attól a helytől, ahol nevelkedett. A csapatban csak olyan párt választanak, akit nem ismernek a régebbi helyükről.[201]
Vannak fajok, ahol az előző évi vagy még régebbi költésből származó fiatalok, tipikusan a hímek még a szüleikkel maradnak, és segítenek felnevelni testvéreiket.[202] A nőstények inkább elhagyják a környéket, és máshol alapítanak családot, vagy segítenek be még egy-két költésbe.
Ökológiai jelentőségük
[szerkesztés]
A madarak ökológiai szerepe igen sokféle.[146] Számos fajuk generalista, és világszerte elterjedt, sokuk pedig szélsőségesen specializálódott az élőhelyéhez vagy a táplálékához. Még egy olyan „ egyszerű” élőhelyen is, mint az erdő, az egyes ökológiai fülkék madárvilága rendkívül eltérő, változatos lehet. Más fajok élnek például az erdő legfelső lombkoronaszintjén, a cserjeszinten, és az aljnövényzet szintjén.
A madarak között egyaránt találhatunk növényevőket, ragadozókat, és dögevőket. A magevők fontos szerepet játszanak az egyes növények elterjesztésében, ugyanis ürülékükkel nagyobb területre hordják szét a magokat.[203] A nektárral táplálkozó madarak a táplálékul szolgáló növény megporzásában játszanak nélkülözhetetlen szerepet (főleg a trópusokon). Ezek között gyakran nagyon szoros koevolúciós kapcsolat figyelhető meg:[204] a növényt kizárólag egyetlen madárfaj képes megporozni, amely pedig csak a növény nektárjával táplálkozik.[205] Ilyen kapcsolat figyelhető meg például számos kolibrifaj esetében.
A ragadozók főleg rovarokra, kisebb emlősökre, más madarakra vadásznak. A vízimadarak a prédaállatok populációinak szabályozásán kívül fontos szerepet játszanak a vízpartok talajának,[206] és a tengervíz tápanyatartalmának[207] megújításában. Ez különösen a tengerpartok, tengeri szigetek tápanyagszegény talaján kolóniákban fészkelő fajoknál fontos.
A partvidéktől távol eső szigeteken gyakran madarak töltik be az egyébként emlősök által elfoglalt ökológiai fülkéket, mivel az emlősök oda nem voltak képesek eljutni. Tipikus példa erre az Új-Zéland szigetén őshonos kereru, vagy a mára már kihalt moák.[203] Új-Zélandon a növényeken olyan adaptációk láthatók, amelyek a moáktól védik őket.[208]
Kapcsolat az emberrel
[szerkesztés]
Mivel a madarak jól láthatók és gyakoriak, az ember már kialakulása idején kapcsolatba került velük.[209] Néha ez a kapcsolat mindkét fél számra előnyös volt, néha hátrány származott belőle valamelyik félnek. A mézkalauzfélék segítik az embereket, hogy a vadméhektől mézet és viaszt szerezzenek, amiért cserébe az emberek részt adnak a zsákmányból.[210] Vannak fajok, például a házi veréb, amelyek az ember szemetén élnek.[211] Néhány fajt elterjedt mezőgazdasági kártevőként tartanak számon.[212] Az ember ugyan a madarak nyomán emelkedett a levegőbe, de néha madarak repülnek a repülők hajtóműveibe, ami a madár halálát és a repülő lezuhanását okozza.[213] A közutak, a gyomirtók, és az ólommérgezés számottevő veszteséget jelentenek a madarak számára. Emellett az élőhelyek beszűkülése és a vadászat is hozzájárul egyes fajok kihalásához. A szélerőművek is pusztítják a madarakat,[214] a kutyák és a macskák is vadásznak rájuk.
A madarak által terjesztett betegségek közé tartozik az influenza (madárinfluenza), a nyugat-nílusi láz szúnyogcsípés révén, a papagájkór, a szalmonella, a campylobacteriosis, a giardiasis és a cryptosporidiosis. Ezek egy részét az emberek is elkaphatják.[215]
Gazdasági jelentőség
[szerkesztés]A gazdasági haszonállatként tartott baromfik húst, tojást és tollat adnak. Az ember belőlük viszi be a legtöbb állati eredetű fehérjét: 2003-ban 76 millió tonna húst és 61 millió tonna tojást.[216] A leggyakoribb faj a házi tyúk, de a pulyka, házi kacsa, lúd is viszonylag gyakori. A vadászatot a kevéssé fejlett területek kivételével általában szórakozásból művelik; a hús melléktermék. Így fácán, fogoly, fajdformák, szalonka, vadpulyka, galambfélék, és Scolopax fajok kerülnek tányérra. Észak- és Dél-Amerikában a vízi szárnyasok vadászata a legfontosabb.[217] Habár ma már a mezőgazdaság segít fenntarthatóvá tenni a vadászatot, régebben sok fajt az ember vadászattal irtott ki, vagy sodort a kihalás szélére.[218]

A tollat hőszigetelésre használják ágyneműkhöz és ruházathoz. A tengeri madarak ürüléke a guanó, amivel trágyáznak, mivel sok nitrogént és foszfort tartalmaz. Chile Bolíviával és Peruval háborúzott is 1879-től 1883-ig az értékes nyersanyagforrásért.[219]
Az ember díszállatként is tart madarakat. Régebben ez több fajt is veszélybe sodort; ma már a befogás illegális, de még mindig előfordul.[220] Sólymokkal és kormoránokkal vadásztak, halásztak. Időszámításunk kezdete körültől ismert, hogy galambokat használtak levelek kézbesítéséhez. A galambposta egészen a második világháború végéig fontos maradt. Ma már a galambászatot turizmus céljából vagy hobbiból űzik,[221] és versenyeken mérik össze galambjaik sebességét.
Az amatőr madármegfigyelők, fényképészek számát milliókban mérik.[222] Sokan télen etetik a környékükön élő madarakat, és az etetők különféle fajokat vonzanak. A madáretetést egy sok milliós üzletág támogatja. Nagy-Britanniában az otthonok 75%-a nyújtott ennivalót a madaraknak a tél egy szakaszában.[223]
Kultúra
[szerkesztés]Vallás
[szerkesztés]Egyes vallásokban a madarak az istenek hírnökei. Makemake a Húsvét-sziget főistene volt, akinek nevében a Tangata manu győztesei lettek a főnökök.[224] Két holló, Hugin és Munin Odin fülébe súgták a híreket.[225] Itália ókori civilizációiban, köztük az etruszk és a római vallásban a papok a madarak viselkedéséből, röptéből jósoltak.[226] Egyes kultúrákban istenítették vagy istenítik őket, mint a pávát Földanya megtestesítőjeként a dravidák.[227] Az inka és a tiwanaku kultúrában a madarak át tudták lépni a határt a földi világ és az alvilág között. A Közép-Andokban legendák szólnak a metafizikai világból jövő és oda távozó madarakról.[228] A Bibliában a galamb jelképként szerepel (héber יוֹנָה).[229]
Képzőművészetek
[szerkesztés]Madárábrázolások már a történelem előtt feltűntek. Láthatók madarak a barlangrajzokon is.[230] Egyes madarak szörnyekként szerepelnek, így a ruk madár vagy a maoriknál a legendás Pouākai, egy nagy, emberevő madár.[231] A későbbi időkben a hatalom szimbólumaivá váltak, a perzsa uralkodó és a mogulok trónszéke pávát ábrázol.[232] A tudományos érdeklődés fellendülésével könyvekhez készültek festmények. Az egyik leghíresebb festő John James Audubon volt, akinek Észak-Amerika madarairól készült képei népszerűvé váltak Európában. Róla nevezték ez a Nemzeti Audubon Társaságot.[233]
Költészet
[szerkesztés]A költészetben is fontos figurák; például az Odüsszeiá-ban csalogányok szerepelnek, Catullusnál egy kismadár szerelmi szimbólummá vált.[234] Az albatrosz és a matróz közötti kapcsolat áll a Samuel Taylor Coleridge: The Rime of the Ancient Mariner című költemény középpontjában. Ebből a versből vált az albatrosz a nehézségek jelképévé.[235] Az angol nyelvben a vulture (keselyű) szóval utalnak azokra az alapokra és befektetőkre, amelyek adósságot vásárolnak olcsón.[236]
A madarakhoz kapcsolódó képzettársítások kultúránként különböznek. A baglyok balszerencsét, boszorkányságot jelentenek, és Afrika és Európa egy részén a halál hírnökei,[237] de Európában bölcsnek tekintik.[238] Az ókori Egyiptomban a búbos banka szent állat volt, és az erény szimbóluma Perzsiában, de Európában tolvajnak vélték, és Skandináviában a háború hírnökeként gondoltak rá.[239]
Zene
[szerkesztés]A zenében a madarak éneke insiprálta a szerzőket. Többen utánozták is, mint Vivaldi és Beethoven, és későbbi szerzők. Amikor a technológia lehetővé tette, akkor a madáréneket felvételről szerkesztették a zenébe. Elsőként Ottorino Respighi tett így. Beatrice Harrison és David Rothenberg duettet énekelnek a madarakkal.[240][241][242]
Rendszerezés
[szerkesztés]
A ma élő madarak két alosztályra tagozódnak, osztályozásuk azonban több helyen is vitatott. Sibley és Ahlquist Phylogeny and Classification of Birds (1990) könyve alapmű,[243] de gyakran vitatják és módosítják. A legtöbb bizonyíték szerint a rendek pontosan megállapíthatók,[244] de a rendek közötti rokonságban nincs egyetértés. Az anatómia, a DNS és a fosszíliák vizsgálatával azonban egyre pontosabb képet kapnak erről is.[245][246]
Futómadár-szabásúak
[szerkesztés]A futómadár-szabásúak (Paleognathae) szájpadjának ősi jellegű, erős és nehéz csontozata van. Szegycsontjuk általában nem tarajos, röpképtelenek vagy csak ritkán repülnek. Két ismert rendjük:
- Struccalakúak rendje (Struthioniformes) – 14 faj
- Tinamualakúak rendje (Tinamiformes) – 47 faj
Újmadárszabásúak
[szerkesztés]Az újmadárszabásúak (Neognathae) csoportjában a szájpad nehéz, csontos váza visszafejlődött, szegycsontjukon nagy csonttaréj biztosítja a mellizmok tapadását. Többségük igen jól repül. Harmincegy rendjük ismert:
- Pingvinalakúak rendje (Sphenisciformes) – 17 faj
- Búváralakúak rendje (Gaviiformes) – 5 faj
- Vöcsökalakúak rendje (Podicipediformes) – 22 faj
- Viharmadár-alakúak rendje (Procellariiformes) – 115 faj
- Gödényalakúak rendje (Pelecaniformes) – 67 faj
- Trópusimadár-alakúak (Phaethontiformes) – 3 faj
- Szulaalakúak (Suliformes)
- Gólyaalakúak rendje (Ciconiiformes) – 127 faj
- Flamingóalakúak rendje (Phoenicopteriformes) – 5 faj
- Lúdalakúak rendje (Anseriformes) – 165 faj
- Sólyomalakúak rendje (Falconiformes) – 303 faj
- Vágómadár-alakúak (Accipitriformes)
- Tyúkalakúak rendje (Galliformes) – 290 faj
- Darualakúak rendje (Gruiformes) – 200 faj
- Guvatgém-alakúak (Eurypygiformes) – 2 faj
- Lilealakúak rendje (Charadriiformes) – 367 faj
- Pusztaityúk-alakúak rendje (Pteroclidiformes) – 16 faj
- Galambalakúak rendje (Columbiformes) – 314 faj
- Papagájalakúak rendje (Psittaciformes) – 361 faj
- Kakukkalakúak rendje (Cuculiformes) – 148 faj
- Turákóalakúak (Musophagiformes)
- Bagolyalakúak rendje (Strigiformes) – 183 faj
- Lappantyúalakúak rendje (Caprimulgiformes) – 120 faj
- Sarlósfecske-alakúak rendje (Apodiformes) – 429 faj
- Egérmadár-alakúak rendje (Coliiformes) – 6 faj
- Trogonalakúak rendje (Trogoniformes) – 39 faj
- Szalakótaalakúak rendje (Coraciiformes) – 214 faj
- Kurolalakúak (Leptosomiformes) – 1 faj
- Szarvascsőrűmadár-alakúak (Bucerotiformes)
- Harkályalakúak rendje (Piciformes) – 405 faj
- Verébalakúak vagy énekesmadár-alakúak rendje (Passeriformes) – 5824 faj
A már nem, vagy ritkán használt rendek
[szerkesztés]- Nandualakúak rendje (Rheiformes) – már nem számít önálló rendnek, lásd a struccalakúak rendjében a nanduféléket.
- Kazuáralakúak rendje (Casuariformes) – már nem számít önálló rendnek, lásd a struccalakúak rendjében a kazuárféléket.
- Kivialakúak rendje (Apterygiformes) – már nem számít önálló rendnek, lásd a struccalakúak rendjében a kiviféléket.
- Sirályalakúak rendje (Lariformes) – már nem számít önálló rendnek, lásd a lilealakúak rendjében a sirályféléket.
- Guvatfürjalakúak rendje (Turniciformes) – már nem számít önálló rendnek, lásd a lilealakúak rendjében a guvatfürjféléket.
- Turákóalakúak rendje (Musophagiformes) – 23 fajt tartalmaz a lista – egyes rendszerezések a kakukkalakúak rendjébe sorolják őket.
- Bankaalakúak rendje (Upupiformes) – Egyes rendszerezések a szalakótaalakúak rendjébe sorolják őket.
- Orrszarvúmadár-alakúak rendje (Bucerotiformes) – Egyes rendszerezések a szalakótaalakúak rendjébe sorolják őket.
Közismert madárnevek
[szerkesztés]Az alábbiakban néhány közismert madártípus neve olvasható (egyszavas madárnevek). A madarak nemét a hím és a tojó elnevezéssel különböztetjük meg. Néhány madártípus hímjének külön elnevezése van, amit zárójelben adunk meg.
Veszélyeztetettség
[szerkesztés]
Habár az ember tevékenysége elősegítette néhány faj terjedését, mint például füsti fecske és seregély, az IUCN Vörös Lista több mint 1200 veszélyeztetett madárfajt tartalmaz.[247][248] Sok faj fogyatkozik. A veszélyeztetettség legtöbbet idézett okai az élőhely elvesztése,[249] a behurcolt fajok ragadozó tevékenysége, balesetek (járművekkel, épületekkel, szélerőművekkel),[250] olajfoltok és növényvédő szerek,[251] behurcolt fajok,[252] vadászat és halászat, valamint a klímaváltozás . A történelmi időkben több mint száz madárfaj halt ki a gyarmatosítás előtt,[253] és a legnagyobb kihalást Melanézia, Polinézia és Mikronézia gyarmatosítása okozta, 750–1800 faj kipusztításával.[254]
Természetvédelmi és kormányzati szervek, csoportok működnek együtt a fajmegőrzésben. Ennek fő eszköze fajok védetté nyilvánítása, élőhelyek védelme, tenyésztés és visszatelepítés. Ezzel 1994 és 2004 között 16 madárfajt sikerült megmenteni a kihalástól, köztük a kaliforniai kondort és a norfolki kecskepapagájt.[255]
Jegyzetek
[szerkesztés]- ↑ http://www.ma.hu/tudomany/174896/Az_Archaeopteryx_madar_volt
- ↑ Brands, Sheila: Systema Naturae 2000 / Classification, Class Aves. Project: The Taxonomicon, 2008. augusztus 14. (Hozzáférés: 2012. június 11.)
- ↑ del Hoyo, Josep. Handbook of Birds of the World, Volume 1: Ostrich to Ducks. Barcelona: Lynx Edicions (1992). ISBN 84-87334-10-5
- ↑ Linnaeus, Carolus. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiae. (Laurentii Salvii), 824. o. (1758)
- ↑ a b c Bradley C. Livezey , RL Zusi (2007. január 1.). „Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion”. Zoological Journal of the Linnean Society 149 (1), 1–95. o. DOI:10.1111/j.1096-3642.2006.00293.x. ISSN 0024-4082. PMID 18784798. PMC 2517308.
- ↑ Padian, Kevin.szerk.: Philip J. Currie and Kevin Padian (eds.): Bird Origins, Encyclopedia of Dinosaurs. San Diego: Academic Press, 41–96. o. (1997). ISBN 0-12-226810-5
- ↑ Gauthier, Jacques.szerk.: Kevin Padian: Saurischian Monophyly and the origin of birds, The Origin of Birds and the Evolution of Flight, Memoirs of the California Academy of Science 8. San Francisco, CA: Published by California Academy of Sciences, 1–55. o. (1986). ISBN 0-940228-14-9
- ↑ a b Gauthier, J., and de Queiroz, K. (2001). "Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name Aves." Pp. 7–41 in New perspectives on the origin and early evolution of birds: proceedings of the International Symposium in Honor of John H. Ostrom (J. A. Gauthier and L. F. Gall, eds.). Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
- ↑ a b (2013) „A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds”. Nature 498 (7454), 359–62. o. DOI:10.1038/nature12168. PMID 23719374.
- ↑ Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.) (2004). The Dinosauria, Second Edition. University of California Press., 861 pp.
- ↑ Senter, P (2007). „A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)”. Journal of Systematic Palaeontology 5, 429–463. o. DOI:10.1017/S1477201907002143.
- ↑ Gauthier, J. (1986). "Saurischian monophyly and the origin of birds." In: K. Padian, ed. The origin of birds and the evolution of flight. San Francisco: California, Acad.Sci. pp.1–55. (Mem.Calif.Acad.Sci.8.)
- ↑ a b Andrea Cau, Tom Brougham, Darren Naish (2015). „The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc(Dinosauria, Maniraptora): Dromaeosaurid or flightless bird?”. PeerJ 3, e1032. o. DOI:10.7717/peerj.1032. PMID 26157616. PMC 4476167.
- ↑ (2011) „Evolving Perceptions on the Antiquity of the Modern Avian Tree, in Living Dinosaurs”. The Evolutionary History of Modern Birds, 306–324. o, Kiadó: John Wiley & Sons LtD. DOI:10.1002/9781119990475.ch12.
- ↑ Influence of Earth's history on the dawn of modern birds. www.sciencedaily.com . American Museum of Natural History, 2015. december 11. (Hozzáférés: 2015. december 11.)
- ↑ a b Michael SY Lee, Andrea Cau, Naish Darren, Dyke Gareth J. (May 2014). „Morphological Clocks in Paleontology, and a Mid-Cretaceous Origin of Crown Aves”. Systematic Biology 63 (1), 442–449. o, Kiadó: Oxford Journals. DOI:10.1093/sysbio/syt110. PMID 24449041.
- ↑ P. D. Alonso, A. C. Milner, R. A. Ketcham, M. J. Cookson, T. B. Rowe (2004). „The avian nature of the brain and inner ear of Archaeopteryx”. Nature 430 (7000), 666–669. o. DOI:10.1038/nature02706. PMID 15295597. PDF fulltext Supplementary info
- ↑ Prum, Richard O. Prum (2008. december 19.). „Who's Your Daddy?”. Science 322 (5909), 1799–1800. o. DOI:10.1126/science.1168808. PMID 19095929.
- ↑ Paul, Gregory S.. Looking for the True Bird Ancestor, Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press, 171–224. o. (2002). ISBN 0-8018-6763-0
- ↑ Mark Norell, Ellison Mick. Unearthing the Dragon: The Great Feathered Dinosaur Discovery. New York: Pi Press (2005). ISBN 0-13-186266-9
- ↑ Borenstein, Seth. „Study traces dinosaur evolution into early birds”, 2014. július 31.. [2014. augusztus 8-i dátummal az eredetiből archiválva] (Hozzáférés: 2017. február 27.)
- ↑ Michael S. Y. Lee, Andrea Cau, Darren Naish, Gareth J. Dyke (2014. augusztus 1.). „Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds”. Science 345 (6196), 562–566. o. DOI:10.1126/science.1252243. PMID 25082702.
- ↑ (2011. július 28.) „An Archaeopteryx-like theropod from China and the origin of Avialae”. Nature 475 (7357), 465–470. o. DOI:10.1038/nature10288. PMID 21796204.
- ↑ Alan H. Turner, D. Pol, J.A. Clarke, G.M. Erickson, M.A. Norell (2007. szeptember 7.). „A basal dromaeosaurid and size evolution preceding avian flight” (PDF). Science 317 (5843), 1378–1381. o. DOI:10.1126/science.1144066. PMID 17823350.
- ↑ X. Xu, Z. Zhou, X. Wang, X. Kuang, F. Zhang, X. Du (2003. január 23.). „Four-winged dinosaurs from China”. Nature 421 (6921), 335–340. o. DOI:10.1038/nature01342. PMID 12540892.
- ↑ Luiggi, Christina: On the Origin of Birds. The Scientist, 2011. július 1. [2012. június 16-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. június 11.)
- ↑ G. Mayr, B. Pohl, S. Hartman, D.S. Peters (2007. január 1.). „The tenth skeletal specimen of Archaeopteryx”. Zoological Journal of the Linnean Society 149 (1), 97–116. o. DOI:10.1111/j.1096-3642.2006.00245.x.
- ↑ a b Zheng, X., et al. (2013. március 15.). „Hind Wings in Basal Birds and the Evolution of Leg Feathers”. Science 339 (6125), 1309–1312. o. DOI:10.1126/science.1228753. PMID 23493711.
- ↑ a b c d Chiappe, Luis M.. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: University of New South Wales Press (2007). ISBN 978-0-86840-413-4
- ↑ Agency France-Presse: Birds survived dino extinction with keen senses. Cosmos Magazine, 2011. április 1. [2015. április 2-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. június 11.)
- ↑ Wang, et al. (2015). „The oldest record of ornithuromorpha from the early cretaceous of China”. Nature Communications 6. DOI:10.1038/ncomms7987. PMID 25942493.
- ↑ Clarke, Julia A. (2004). „Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae)” (PDF). Bulletin of the American Museum of Natural History 286, 1–179. o. DOI:<0001:MPTASO>2.0.CO;2 10.1206/0003-0090(2004)286<0001:MPTASO>2.0.CO;2. (Hozzáférés: 2017. március 1.)
- ↑ Clarke, Julia A. (2004). „Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae)” (PDF). Bulletin of the American Museum of Natural History 286, 1–179. o. DOI:<0001:MPTASO>2.0.CO;2 10.1206/0003-0090(2004)286<0001:MPTASO>2.0.CO;2. (Hozzáférés: 2017. március 1.)
- ↑ A. Louchart, L. Viriot (2011). „From snout to beak: the loss of teeth in birds”. Trends In Ecology & Evolution 26 (12), 663–673. o. [2014. július 28-i dátummal az eredetiből archiválva]. DOI:10.1016/j.tree.2011.09.004. (Hozzáférés: 2017. március 1.)
- ↑ J. A. Clarke, Z. Zhou, F. Zhang (2006. március 1.). „Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui”. Journal of Anatomy 208 (3), 287–308. o. DOI:10.1111/j.1469-7580.2006.00534.x. PMID 16533313. PMC 2100246.
- ↑ Ritchison, Gary: Bird biogeography. Avian Biology. Eastern Kentucky University. (Hozzáférés: 2008. április 10.)
- ↑ Clements, James F.. The Clements Checklist of Birds of the World, 6th, Ithaca: Cornell University Press (2007). ISBN 978-0-8014-4501-9
- ↑ Gill, Frank. Birds of the World: Recommended English Names. Princeton: Princeton University Press (2006). ISBN 978-0-691-12827-6
- ↑ Clarke, Julia A. (2005). „Definitive fossil evidence for the extant avian radiation in the Cretaceous” (PDF). Nature 433 (7023), 305–308. o. DOI:10.1038/nature03150. PMID 15662422. Nature.com, Supporting information
- ↑ Clarke, J.A. (2004). „Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae)”. Bulletin of the American Museum of Natural History 286, 1–179. o. [2015. június 19-i dátummal az eredetiből archiválva]. DOI:<0001:mptaso>2.0.co;2 10.1206/0003-0090(2004)286<0001:mptaso>2.0.co;2. (Hozzáférés: 2017. március 2.)
- ↑ a b Ericson, Per G.P., et al. (2006). „Diversification of Neoaves: integration of molecular sequence data and fossils” (PDF). Biology Letters 2 (4), 543–547. o. [2009. március 25-i dátummal az eredetiből archiválva]. DOI:10.1098/rsbl.2006.0523. PMID 17148284. (Hozzáférés: 2017. március 2.)
- ↑ Joseph W. Brown, R.B. Payne, D.P. Mindell (2007. június 1.). „Nuclear DNA does not reconcile 'rocks' and 'clocks' in Neoaves: a comment on Ericson et al.”. Biology Letters 3 (3), 257–259. o. DOI:10.1098/rsbl.2006.0611. PMID 17389215. PMC 2464679.
- ↑ Ornithologists Publish Most Comprehensive Avian Tree of Life
- ↑ Newton, Ian. The Speciation and Biogeography of Birds. Amsterdam: Academic Press, 463. o. (2003). ISBN 0-12-517375-X
- ↑ a b Brooke, Michael. Albatrosses And Petrels Across The World. Oxford: Oxford University Press (2004). ISBN 0-19-850125-0
- ↑ Schreiber, Elizabeth Anne. Biology of Marine Birds. Boca Raton: CRC Press (2001). ISBN 0-8493-9882-7
- ↑ Sato, Katsufumi, et al. (2002. május 1.). „Buoyancy and maximal diving depth in penguins: do they control inhaling air volume?”. Journal of Experimental Biology 205 (9), 1189–1197. o. PMID 11948196.
- ↑ Hill, David. The Pheasant: Ecology, Management, and Conservation. Oxford: BSP Professional (1988). ISBN 0-632-02011-3
- ↑ Spreyer, Mark F.: Monk Parakeet (Myiopsitta monachus). The Birds of North America. Cornell Lab of Ornithology, 1998. DOI:10.2173/bna.322. (Hozzáférés: 2015. december 13.)
- ↑ Arendt, Wayne J. (1988. január 1.). „Range Expansion of the Cattle Egret, (Bubulcus ibis) in the Greater Caribbean Basin”. Colonial Waterbirds 11 (2), 252–62. o. DOI:10.2307/1521007. JSTOR 1521007.
- ↑ Bierregaard, R.O..szerk.: Josep del Hoyo, Andrew Elliott and Jordi Sargatal (eds.): Yellow-headed Caracara, Handbook of the Birds of the World. Volume 2; New World Vultures to Guineafowl. Barcelona: Lynx Edicions (1994). ISBN 84-87334-15-6
- ↑ Juniper, Tony. Parrots: A Guide to the Parrots of the World. London: Christopher Helm (1998). ISBN 0-7136-6933-0
- ↑ a b c d e f g h i j k l m n o p q r s t u v Gill, Frank. Ornithology. New York: WH Freeman and Co (1995). ISBN 0-7167-2415-4
- ↑ Anton Reiner, David J. Perkel, Claudio V. Mello, Erich D. Jarvis: Songbirds and the Revised Avian Brain Nomenclature. In: H. Philip Zeigler, Peter Marler (Hrsg): Behavioral Neurobiology of Birdsong. Annals of the New York Academy of Sciences. Bd. 1016, 2004, S. 77–108, doi:10.1196/annals.1298.013, PMC 2481519
- ↑ Gesamter Absatz nach: Seweryn Olkowicz, Martin Kocourek, Radek K. Lučan, Michal Porteš, W. Tecumseh Fitch, Suzana Herculano-Houzel, Pavel Němec: Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences of the United States of America. Bd. 113, Nr. 26, 2016, S. 7255–7260, doi:10.1073/pnas.1517131113
- ↑ Wilkie, Susan E., et al. (1998. február 1.). „The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus)”. Biochemical Journal 330 (Pt 1), 541–47. o. PMID 9461554. PMC 1219171.
- ↑ Féányérzékeny sejtek az agyban[halott link]
- ↑ Háziállat.hu a baglyokról
- ↑ Sales, James (2005). „The endangered kiwi: a review” (PDF). Folia Zoologica 54 (1–2), 1–20. o. [2007. szeptember 26-i dátummal az eredetiből archiválva]. (Hozzáférés: 2017. március 6.)
- ↑ Ehrlich, Paul R.: The Avian Sense of Smell. Birds of Stanford. Stanford University, 1988. (Hozzáférés: 2007. december 13.)
- ↑ Lequette, Benoit (1989. augusztus 1.). „Olfaction in Subantarctic seabirds: Its phylogenetic and ecological significance” (PDF). The Condor 91 (3), 732–35. o. [2013. február 23-i dátummal az eredetiből archiválva]. DOI:10.2307/1368131.
- ↑ Christine Maira Hein, Svenja Engels, Dmitry Kishkinev, Henrik Mouritsen: Robins have a magnetic compass in both eyes. In: Nature. Bd. 471, E1, 2011, doi:10.1038/nature09875.
- ↑ Ehrlich, Paul R.: Adaptations for Flight. Birds of Stanford. Stanford University, 1988. (Hozzáférés: 2007. december 13.) Based on The Birder's Handbook (Paul Ehrlich, David Dobkin, and Darryl Wheye. 1988. Simon and Schuster, New York.)
- ↑ Noll, Paul. „The Avian Skeleton”, paulnoll.com (Hozzáférés: 2007. december 13.)
- ↑ „Skeleton of a typical bird”, Fernbank Science Center's Ornithology Web (Hozzáférés: 2007. december 13.)
- ↑ Demay, Ida S. (1940). „A Study of the Pterylosis and Pneumaticity of the Screamer”. The Condor 42 (2), 112–118. o. DOI:10.2307/1364475. JSTOR 1364475.
- ↑ Guthrie, R. Dale: How We Use and Show Our Social Organs. Body Hot Spots: The Anatomy of Human Social Organs and Behavior. [2007. június 21-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. október 19.)
- ↑ Belthoff, James R. (1994. augusztus 1.). „Plumage Variation, Plasma Steroids and Social Dominance in Male House Finches”. The Condor 96 (3), 614–25. o. DOI:10.2307/1369464.
- ↑ a b c Pettingill Jr. OS. Ornithology in Laboratory and Field. Burgess Publishing Co (1970). ISBN 0-12-552455-2
- ↑ de Beer SJ, Lockwood GM, Raijmakers JHFS, Raijmakers JMH, Scott WA, Oschadleus HD, Underhill LG (2001). "SAFRING Bird Ringing Manual Archiválva 2017. október 19-i dátummal a Wayback Machine-ben".
- ↑ Gargallo, Gabriel (1994. június 1.). „Flight Feather Moult in the Red-Necked Nightjar Caprimulgus ruficollis”. Journal of Avian Biology 25 (2), 119–24. o. DOI:10.2307/3677029. JSTOR 3677029.
- ↑ Mayr, Ernst (1954). „The tail molt of small owls” (PDF). The Auk 71 (2), 172–78. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/4081571.
- ↑ Payne, Robert B: Birds of the World, Biology 532. Bird Division, University of Michigan Museum of Zoology. [2012. február 26-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. október 20.)
- ↑ Turner, J. Scott (1997). „On the thermal capacity of a bird's egg warmed by a brood patch”. Physiological Zoology 70 (4), 470–80. o. DOI:10.1086/515854. PMID 9237308.
- ↑ Walther, Bruno A. (2005). „Elaborate ornaments are costly to maintain: evidence for high maintenance handicaps”. Behavioural Ecology 16 (1), 89–95. o. DOI:10.1093/beheco/arh135.
- ↑ Matthew D. Shawkey, R. Pillai, Geoffrey E. Hill (2003). „Chemical warfare? Effects of uropygial oil on feather-degrading bacteria”. Journal of Avian Biology 34 (4), 345–49. o. DOI:10.1111/j.0908-8857.2003.03193.x.
- ↑ Ehrlich, Paul R. (1986). „The Adaptive Significance of Anting” (PDF). The Auk 103 (4), 835. o. [2013. május 24-i dátummal az eredetiből archiválva].
- ↑ Lucas, Alfred M.. Avian Anatomy—integument. East Lansing, Michigan, US: USDA Avian Anatomy Project, Michigan State University, 67, 344, 394–601. o. (1972)
- ↑ Balgooyen, Thomas G. (1971. október 1.). „Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius)” (PDF). Condor 73 (3), 382–85. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/1365774. JSTOR 1365774.
- ↑ a b c Attenborough, David. The Life of Birds. Princeton: Princeton University Press (1998). ISBN 0-691-01633-X
- ↑ a b Battley, Phil F., et al. (2000. január 1.). „Empirical evidence for differential organ reductions during trans-oceanic bird flight”. Proceedings of the Royal Society B 267 (1439), 191–5. o. DOI:10.1098/rspb.2000.0986. PMID 10687826. PMC 1690512. (Erratum in Proceedings of the Royal Society B 267(1461):2567.)
- ↑ a b Maina, John N. (2006. november 1.). „Development, structure, and function of a novel respiratory organ, the lung-air sac system of birds: to go where no other vertebrate has gone”. Biological Reviews 81 (4), 545–79. o. DOI:10.1017/S1464793106007111. PMID 17038201.
- ↑ a b Suthers, Roderick A. (2004. június 1.). „Producing song: the vocal apparatus”. Ann. N. Y. Acad. Sci. 1016, 109–29. o. DOI:10.1196/annals.1298.041. PMID 15313772.
- ↑ Fitch, W. T. (1999). „Acoustic exaggeration of size in birds via tracheal elongation: comparative and theoretical analyses”. Journal of Zoology 248, 31–48. o. DOI:10.1017/S095283699900504X.
- ↑ Whittow, G. (2000). Sturkie's Avian Physiology/ edited by G. Causey Whittow. San Diego : Academic Press, 2000.
- ↑ a b Hoagstrom, C.W. (2002). Vertebrate Circulation. Magill's Encyclopedia of Science: Animal Life. Vol 1, pp 217-219. Pasadena, California, Salem Press.
- ↑ a b Hill, Richard W. (2012) Animal Physiology/ Richard W. Hill, Gordon A. Wyse, Margaret Anderson. Third Edition pp 647-678. Sinauer Associates, 23 Plumtree Road, Sunderland, MA 01375 USA
- ↑ Ehrlich, Paul R.: Drinking. Birds of Stanford. Stanford University, 1988. (Hozzáférés: 2007. december 13.)
- ↑ Tsahar, Ella, et al. (2005). „Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores”. Journal of Experimental Biology 208 (6), 1025–34. o. DOI:10.1242/jeb.01495. PMID 15767304.
- ↑ Skadhauge, E, et al. (2003). „Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?”. Comparative Biochemistry and Physiology A 134 (4), 749–755. o. DOI:10.1016/S1095-6433(03)00006-0. PMID 12814783.
- ↑ Marion R., Carol A. Beuchat (1997. április 1.). „Ammonia excretion by hummingbirds”. Nature 386 (6625), 561–62. o. DOI:10.1038/386561a0.
- ↑ Mora, J., et al. (1965). „The regulation of urea-biosynthesis enzymes in vertebrates”. Biochemical Journal 96 (1), 28–35. o. PMID 14343146. PMC 1206904.
- ↑ Packard, Gary C. (1966). „The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates”. The American Naturalist 100 (916), 667–82. o. DOI:10.1086/282459. JSTOR 2459303.
- ↑ A. Göth, D.T. Booth (2005. március 1.). „Temperature-dependent sex ratio in a bird”. Biology Letters 1, 31–3. o. DOI:10.1098/rsbl.2004.0247. PMID 17148121. PMC 1629050.
- ↑ Yong, Ed: Phenomena: Not Exactly Rocket Science How Chickens Lost Their Penises (And Ducks Kept Theirs). Phenomena.nationalgeographic.com. (Hozzáférés: 2013. október 3.)
- ↑ Ornithology, 3rd Edition - Waterfowl: Order Anseriformes. [2015. június 22-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. október 3.)
- ↑ McCracken, KG (2000). „The 20-cm Spiny Penis of the Argentine Lake Duck (Oxyura vittata)”. The Auk 117 (3), 820–825. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:[0820:TCSPOT2.0.CO;2 10.1642/0004-8038(2000)117[0820:TCSPOT]2.0.CO;2].
- ↑ Gionfriddo, James P. (1995. február 1.). „Grit Use by House Sparrows: Effects of Diet and Grit Size” (PDF). Condor 97 (1), 57–67. o. DOI:10.2307/1368983.
- ↑ Warham, John (1977. május 1.). „The incidence, function and ecological significance of petrel stomach oils” (PDF). Proceedings of the New Zealand Ecological Society 24 (3), 84–93. o.
- ↑ Dumbacher, J.P., et al. (1992. október 1.). „Homobatrachotoxin in the genus Pitohui: chemical defense in birds?”. Science 258 (5083), 799–801. o. DOI:10.1126/science.1439786. PMID 1439786.
- ↑ N. R. Longrich, S. L. Olson (2011. január 5.). „The bizarre wing of the Jamaican flightless ibis Xenicibis xympithecus: a unique vertebrate adaptation”. Proceedings of the Royal Society B: Biological Sciences 278 (1716), 2333–2337. o. DOI:10.1098/rspb.2010.2117. (Hozzáférés: 2015. november 12.)
- ↑ Michel Robert, Raymond McNeil, Alain Leduc (1989. január 1.). „Conditions and significance of night feeding in shorebirds and other water birds in a tropical lagoon” (PDF). The Auk 106 (1), 94–101. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/4087761.
- ↑ McNab, Brian K. (1994. október 1.). „Energy Conservation and the Evolution of Flightlessness in Birds”. The American Naturalist 144 (4), 628–42. o. DOI:10.1086/285697. JSTOR 2462941.
- ↑ Roots, Clive. Flightless Birds. Westport: Greenwood Press (2006). ISBN 978-0-313-33545-7
- ↑ Christopher E. Kovacs , R. A. Meyers (2000). „Anatomy and histochemistry of flight muscles in a wing-propelled diving bird, the Atlantic Puffin, Fratercula arctica”. Journal of Morphology 244 (2), 109–25. o. DOI:<109::AID-JMOR2>3.0.CO;2-0 10.1002/(SICI)1097-4687(200005)244:2<109::AID-JMOR2>3.0.CO;2-0. PMID 10761049.
- ↑ How Do Birds Eat If They Have No Teeth? | Blog | eNature. wild.enature.com . [2016. április 14-i dátummal az eredetiből archiválva]. (Hozzáférés: 2017. március 13.)
- ↑ N Reid: Birds on New England wool properties – A woolgrower guide (PDF). Land, Water & Wool Northern Tablelands Property Fact Sheet. Australian Government – Land and Water Australia, 2006. [2011. március 15-i dátummal az eredetiből archiválva]. (Hozzáférés: 2010. július 17.)
- ↑ D. Paton , C. B. G. Collins (1989. április 1.). „Bills and tongues of nectar-feeding birds: A review of morphology, function, and performance, with intercontinental comparisons”. Australian Journal of Ecology 14 (4), 473–506. o. DOI:10.1111/j.1442-9993.1989.tb01457.x.
- ↑ Baker, Myron Charles (1973. április 1.). „Niche Relationships Among Six Species of Shorebirds on Their Wintering and Breeding Ranges”. Ecological Monographs 43 (2), 193–212. o. DOI:10.2307/1942194. JSTOR 1942194.
- ↑ Cherel, Yves (2002). „Food and feeding ecology of the sympatric thin-billed Pachyptila belcheri and Antarctic P. desolata prions at Iles Kerguelen, Southern Indian Ocean”. Marine Ecology Progress Series 228, 263–81. o. DOI:10.3354/meps228263.
- ↑ Jenkin, Penelope M. (1957). „The Filter-Feeding and Food of Flamingoes (Phoenicopteri)”. Philosophical Transactions of the Royal Society B 240 (674), 401–93. o. DOI:10.1098/rstb.1957.0004. JSTOR 92549.
- ↑ Miyazaki, Masamine (1996. július 1.). „Vegetation cover, kleptoparasitism by diurnal gulls and timing of arrival of nocturnal Rhinoceros Auklets” (PDF). The Auk 113 (3), 698–702. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/3677021. JSTOR 3677021.
- ↑ Bélisle, Marc (1995. augusztus 1.). „Predation and kleptoparasitism by migrating Parasitic Jaegers” (PDF). The Condor 97 (3), 771–781. o. DOI:10.2307/1369185.
- ↑ Vickery, J. A. (1994. május 1.). „The Kleptoparasitic Interactions between Great Frigatebirds and Masked Boobies on Henderson Island, South Pacific” (PDF). The Condor 96 (2), 331–40. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/1369318. JSTOR 1369318.
- ↑ Hiraldo, F.C. (1991). „Unspecialized exploitation of small carcasses by birds”. Bird Studies 38 (3), 200–07. o. DOI:10.1080/00063659109477089.
- ↑ Engel, Sophia Barbara. Racing the wind: Water economy and energy expenditure in avian endurance flight [archivált változat]. University of Groningen (2005). ISBN 90-367-2378-7. Hozzáférés ideje: 2017. március 15. [archiválás ideje: 2020. április 5.]
- ↑ B.I. Tieleman, J.B. Williams (1999). „The role of hyperthermia in the water economy of desert birds”. Physiol. Biochem. Zool. 72 (1), 87–100. o. DOI:10.1086/316640. PMID 9882607.
- ↑ Schmidt-Nielsen, Knut (1960. május 1.). „The Salt-Secreting Gland of Marine Birds”. Circulation 21 (5), 955–967. o. DOI:10.1161/01.CIR.21.5.955.
- ↑ Hallager, Sara L. (1994). „Drinking methods in two species of bustards”. Wilson Bull. 106 (4), 763–764. o.
- ↑ MacLean, Gordon L. (1983. június 1.). „Water Transport by Sandgrouse”. BioScience 33 (6), 365–369. o. DOI:10.2307/1309104. JSTOR 1309104.
- ↑ Eraud C (2008). „The crop milk: a potential new route for carotenoid-mediated parental effects”. Journal of Avian Biology 39 (2), 247–251. o. DOI:10.1111/j.0908-8857.2008.04053.x.
- ↑ Mario, Principato, et al. (2005). „The alterations of plumage of parasitic origin”. Italian Journal of Animal Science 4, 296–299. o. [2013. június 17-i dátummal az eredetiből archiválva]. (Hozzáférés: 2017. március 16.)
- ↑ Hannah C. Revis, Deborah A. Waller (2004). „Bactericidal and fungicidal activity of ant chemicals on feather parasites: an evaluation of anting behavior as a method of self-medication in songbirds”. The Auk 121 (4), 1262–1268. o. DOI:[1262:BAFAOA2.0.CO;2 10.1642/0004-8038(2004)121[1262:BAFAOA]2.0.CO;2].
- ↑ Clayton, Dale H., et al. (2010). „How Birds Combat Ectoparasites”. The Open Ornithology Journal 3, 41–71. o. [2017. augusztus 11-i dátummal az eredetiből archiválva]. DOI:10.2174/1874453201003010041. (Hozzáférés: 2017. március 16.)
- ↑ Klaassen, Marc (1996. január 1.). „Metabolic constraints on long-distance migration in birds”. Journal of Experimental Biology 199 (1), 57–64. o. PMID 9317335.
- ↑ Gill, Frank. Ornithology, 2nd, New York: W.H. Freeman (1995). ISBN 0-7167-2415-4
- ↑ „Long-distance Godwit sets new record”, BirdLife International, 2007. május 4.. [2013. október 2-i dátummal az eredetiből archiválva] (Hozzáférés: 2007. december 13.)
- ↑ Shaffer, Scott A., et al. (2006). „Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer”. Proceedings of the National Academy of Sciences of the United States of America 103 (34), 12799–802. o. DOI:10.1073/pnas.0603715103. PMID 16908846.
- ↑ Croxall, John P., et al. (2005). „Global Circumnavigations: Tracking year-round ranges of nonbreeding Albatrosses”. Science 307 (5707), 249–50. o. DOI:10.1126/science.1106042. PMID 15653503.
- ↑ Wilson, W. Herbert, Jr. (1999). „Bird feeding and irruptions of northern finches:are migrations short stopped?” (PDF). North America Bird Bander 24 (4), 113–21. o. [2013. május 24-i dátummal az eredetiből archiválva].
- ↑ Nilsson, Anna L. K. (2006). „Do partial and regular migrants differ in their responses to weather?”. The Auk 123 (2), 537–47. o. DOI:[537:DPARMD2.0.CO;2 10.1642/0004-8038(2006)123[537:DPARMD]2.0.CO;2].
- ↑ Rabenold, Kerry N. (1985). „Variation in Altitudinal Migration, Winter Segregation, and Site Tenacity in two subspecies of Dark-eyed Juncos in the southern Appalachians” (PDF). The Auk 102 (4), 805–19. o.
- ↑ Collar, Nigel J..szerk.: Josep del Hoyo, Andrew Elliott and Jordi Sargatal (eds.): Family Psittacidae (Parrots), Handbook of the Birds of the World, Volume 4: Sandgrouse to Cuckoos. Barcelona: Lynx Edicions (1997). ISBN 84-87334-22-9
- ↑ Matthews, G. V. T. (1953. szeptember 1.). „Navigation in the Manx Shearwater”. Journal of Experimental Biology 30 (2), 370–96. o.
- ↑ Mouritsen, Henrik (2001. november 15.). „Migrating songbirds tested in computer-controlled Emlen funnels use stellar cues for a time-independent compass”. Journal of Experimental Biology 204 (8), 3855–65. o. PMID 11807103.
- ↑ Deutschlander, Mark E. (1999. április 15.). „The case for light-dependent magnetic orientation in animals”. Journal of Experimental Biology 202 (8), 891–908. o. PMID 10085262.
- ↑ Möller, Anders Pape (1988). „Badge size in the house sparrow Passer domesticus”. Behavioral Ecology and Sociobiology 22 (5), 373–78. o. DOI:10.1007/BF00295107.
- ↑ Thomas, Betsy Trent (1990. augusztus 1.). „Nesting Behavior of Sunbitterns (Eurypyga helias) in Venezuela” (PDF). The Condor 92 (3), 576–81. o. [2013. május 24-i dátummal az eredetiből archiválva]. DOI:10.2307/1368675.
- ↑ Pickering, S. P. C. (2001). „Courtship behaviour of the Wandering Albatross Diomedea exulans at Bird Island, South Georgia” (PDF). Marine Ornithology 29 (1), 29–37. o.
- ↑ Pruett-Jones, S. G. (1990. május 1.). „Sexual Selection Through Female Choice in Lawes' Parotia, A Lek-Mating Bird of Paradise”. Evolution 44 (3), 486–501. o. DOI:10.2307/2409431.
- ↑ Genevois, F. (1994). „Male Blue Petrels reveal their body mass when calling”. Ethology Ecology and Evolution 6 (3), 377–83. o. [2007. december 24-i dátummal az eredetiből archiválva]. DOI:10.1080/08927014.1994.9522988. (Hozzáférés: 2017. március 19.)
- ↑ Jouventin, Pierre (1999. június 1.). „Finding a parent in a king penguin colony: the acoustic system of individual recognition”. Animal Behaviour 57 (6), 1175–83. o. DOI:10.1006/anbe.1999.1086. PMID 10373249.
- ↑ Templeton, Christopher N. (2005). „Allometry of Alarm Calls: Black-Capped Chickadees Encode Information About Predator Size”. Science 308 (5730), 1934–37. o. DOI:10.1126/science.1108841. PMID 15976305.
- ↑ Miskelly, C. M. (1987. július 1.). „The identity of the hakawai”. Notornis 34 (2), 95–116. o.
- ↑ Murphy, Stephen (2003). „The breeding biology of palm cockatoos (Probosciger aterrimus): a case of a slow life history”. Journal of Zoology 261 (4), 327–39. o. DOI:10.1017/S0952836903004175.
- ↑ a b Sekercioglu, Cagan Hakki.szerk.: Josep del Hoyo, Andrew Elliott and David Christie (eds.): Foreword, Handbook of the Birds of the World, Volume 11: Old World Flycatchers to Old World Warblers. Barcelona: Lynx Edicions, 48. o. (2006). ISBN 84-96553-06-X
- ↑ Terborgh, John (2005). „Mixed flocks and polyspecific associations: Costs and benefits of mixed groups to birds and monkeys”. American Journal of Primatology 21 (2), 87–100. o. DOI:10.1002/ajp.1350210203.
- ↑ Hutto, Richard L. (1988. január 1.). „Foraging Behavior Patterns Suggest a Possible Cost Associated with Participation in Mixed-Species Bird Flocks”. Oikos 51 (1), 79–83. o. DOI:10.2307/3565809. JSTOR 3565809.
- ↑ Au, David W. K. (1986. augusztus 1.). „Seabird interactions with Dolphins and Tuna in the Eastern Tropical Pacific” (PDF). The Condor 88 (3), 304–17. o. DOI:10.2307/1368877.
- ↑ Anne, O. (1983. június 1.). „Dwarf mongoose and hornbill mutualism in the Taru desert, Kenya”. Behavioral Ecology and Sociobiology 12 (3), 181–90. o. DOI:10.1007/BF00290770.
- ↑ Gauthier-Clerc, Michael (2000). „Sleep-Vigilance Trade-off in Gadwall during the Winter Period” (PDF). The Condor 102 (2), 307–13. o. [2004. december 27-i dátummal az eredetiből archiválva]. DOI:[0307:SVTOIG2.0.CO;2 10.1650/0010-5422(2000)102[0307:SVTOIG]2.0.CO;2]. JSTOR 1369642.
- ↑ Bäckman, Johan (2002. április 1.). „Harmonic oscillatory orientation relative to the wind in nocturnal roosting flights of the swift Apus apus”. The Journal of Experimental Biology 205 (7), 905–910. o. PMID 11916987.
- ↑ Rattenborg, Niels C. (2006). „Do birds sleep in flight?”. Die Naturwissenschaften 93 (9), 413–25. o. DOI:10.1007/s00114-006-0120-3. PMID 16688436.
- ↑ Milius, S. (1999. február 6.). „Half-asleep birds choose which half dozes”. Science News Online 155 (6), 86. o. DOI:10.2307/4011301. JSTOR 4011301.
- ↑ Beauchamp, Guy (1999). „The evolution of communal roosting in birds: origin and secondary losses”. Behavioural Ecology 10 (6), 675–87. o. DOI:10.1093/beheco/10.6.675.
- ↑ Buttemer, William A. (1985). „Energy relations of winter roost-site utilization by American goldfinches (Carduelis tristis)” (PDF). Oecologia 68 (1), 126–32. o. DOI:10.1007/BF00379484.
- ↑ Buckley, F. G. (1968. január 1.). „Upside-down Resting by Young Green-Rumped Parrotlets (Forpus passerinus)”. The Condor 70 (1), 89. o. DOI:10.2307/1366517.
- ↑ Carpenter, F. Lynn (1974). „Torpor in an Andean Hummingbird: Its Ecological Significance”. Science 183 (4124), 545–47. o. DOI:10.1126/science.183.4124.545. PMID 17773043.
- ↑ McKechnie, Andrew E. (2007). „Torpor in an African caprimulgid, the freckled nightjar Caprimulgus tristigma”. Journal of Avian Biology 38 (3), 261–66. o. DOI:10.1111/j.2007.0908-8857.04116.x.
- ↑ Frith, C.B (1981). „Displays of Count Raggi's Bird-of-Paradise Paradisaea raggiana and congeneric species”. Emu 81 (4), 193–201. o. DOI:10.1071/MU9810193.
- ↑ Freed, Leonard A. (1987). „The Long-Term Pair Bond of Tropical House Wrens: Advantage or Constraint?”. The American Naturalist 130 (4), 507–25. o. DOI:10.1086/284728.
- ↑ Gowaty, Patricia A. (1983). „Male Parental Care and Apparent Monogamy among Eastern Bluebirds (Sialia sialis)”. The American Naturalist 121 (2), 149–60. o. DOI:10.1086/284047.
- ↑ Westneat, David F. (2003). „Extra-pair paternity in birds: Causes, correlates, and conflict”. Annual Review of Ecology, Evolution, and Systematics 34, 365–96. o. DOI:10.1146/annurev.ecolsys.34.011802.132439.[halott link]
- ↑ Gowaty, Patricia A. (1998). „Ultimate causation of aggressive and forced copulation in birds: Female resistance, the CODE hypothesis, and social monogamy”. American Zoologist 38 (1), 207–25. o. DOI:10.1093/icb/38.1.207.
- ↑ T.R. Birkhead, P. Møller (1993). „Sexual selection and the temporal separation of reproductive events: sperm storage data from reptiles, birds and mammals”. Biological Journal of the Linnean Society 50, 295–311. o. DOI:10.1111/j.1095-8312.1993.tb00933.x.
- ↑ Sheldon, B (1994). „Male Phenotype, Fertility, and the Pursuit of Extra-Pair Copulations by Female Birds”. Proceedings of the Royal Society B 257 (1348), 25–30. o. DOI:10.1098/rspb.1994.0089.
- ↑ Wei, G (2005). „Copulations and mate guarding of the Chinese Egret”. Waterbirds 28 (4), 527–30. o. DOI:[527:CAMGOT2.0.CO;2 10.1675/1524-4695(2005)28[527:CAMGOT]2.0.CO;2].
- ↑ Bagemihl, Bruce. Biological exuberance: Animal homosexuality and natural diversity. New York: St. Martin's, 1999. pp. 479–655. One hundred species are described in detail.
- ↑ Short, Lester L.. Birds of the World and their Behavior. New York: Henry Holt and Co (1993). ISBN 0-8050-1952-9
- ↑ Burton, R. Bird Behavior. Alfred A. Knopf, Inc (1985). ISBN 0-394-53957-5
- ↑ Schamel, D (2004). „Mate guarding, copulation strategies and paternity in the sex-role reversed, socially polyandrous red-necked phalarope Phalaropus lobatus” (PDF). Behaviour Ecology and Sociobiology 57 (2), 110–18. o. [2020. március 28-i dátummal az eredetiből archiválva]. DOI:10.1007/s00265-004-0825-2. (Hozzáférés: 2017. március 22.)
- ↑ H. Kokko, M. Harris, S. Wanless (2004). „Competition for breeding sites and site-dependent population regulation in a highly colonial seabird, the common guillemot Uria aalge”. Journal of Animal Ecology 73 (2), 367–76. o. DOI:10.1111/j.0021-8790.2004.00813.x.
- ↑ L. Booker, M. Booker (1991). „Why Are Cuckoos Host Specific?”. Oikos 57 (3), 301–09. o. DOI:10.2307/3565958. JSTOR 3565958.
- ↑ a b Hansell M (2000). Bird Nests and Construction Behaviour. University of Cambridge Press ISBN 0-521-46038-7
- ↑ L. Lafuma, M. Lambrechts, M. Raymond (2001). „Aromatic plants in bird nests as a protection against blood-sucking flying insects?”. Behavioural Processes 56 (2), 113–20. o. DOI:10.1016/S0376-6357(01)00191-7.
- ↑ Warham, J. (1990) The Petrels: Their Ecology and Breeding Systems London: Academic Press ISBN 0-12-735420-4.
- ↑ Jones DN, Dekker, René WRJ, Roselaar, Cees S (1995). The Megapodes. Bird Families of the World 3. Oxford University Press: Oxford. ISBN 0-19-854651-3
- ↑ AnAge: The animal ageing and longevity database. Human Ageing and Genomics Resources. (Hozzáférés: 2014. szeptember 26.)
- ↑ Animal diversity web. University of Michigan, Museum of Zoology. (Hozzáférés: 2014. szeptember 26.)
- ↑ Elliot A (1994). "Family Megapodiidae (Megapodes)" in Handbook of the Birds of the World. Volume 2; New World Vultures to Guineafowl (eds del Hoyo J, Elliott A, Sargatal J) Lynx Edicions:Barcelona. ISBN 84-87334-15-6
- ↑ Metz VG, Schreiber EA (2002). "Great Frigatebird (Fregata minor)" In The Birds of North America, No 681, (Poole, A. and Gill, F., eds) The Birds of North America Inc: Philadelphia
- ↑ Young, Euan. Skua and Penguin. Predator and Prey.. Cambridge University Press, 1994, p. 453.
- ↑ Ekman, J. (2006). „Family living amongst birds”. Journal of Avian Biology 37 (4), 289–98. o. DOI:10.1111/j.2006.0908-8857.03666.x.
- ↑ Cockburn A. Why do so many Australian birds cooperate? Social evolution in the Corvida, Frontiers in Population Ecology. Melbourne: CSIRO, 21–42. o. (1996)
- ↑ Cockburn, Andrew (2006). „Prevalence of different modes of parental care in birds”. Proceedings of the Royal Society B 273 (1592), 1375–83. o. DOI:10.1098/rspb.2005.3458. PMID 16777726. PMC 1560291.
- ↑ Gaston AJ (1994). Ancient Murrelet (Synthliboramphus antiquus). In The Birds of North America, No. 132 (A. Poole and F. Gill, Eds.). Philadelphia: The Academy of Natural Sciences; Washington, D.C.: The American Ornithologists' Union.
- ↑ H.C. Schaefer, et al. (2004). „Life-history of two African Sylvia warblers: low annual fecundity and long post-fledging care”. Ibis 146 (3), 427–37. o. DOI:10.1111/j.1474-919X.2004.00276.x.
- ↑ J.C Alonso, L.M. Bautista, J.A. Alonso (2004). „Family-based territoriality vs flocking in wintering common cranes Grus grus”. Journal of Avian Biology 35 (5), 434–44. o. DOI:10.1111/j.0908-8857.2004.03290.x.
- ↑ Davies N (2000). Cuckoos, Cowbirds and other Cheats. T. & A. D. Poyser: London ISBN 0-85661-135-2
- ↑ Sorenson, M. (1997). „Effects of intra- and interspecific brood parasitism on a precocial host, the canvasback, Aythya valisineria”. Behavioral Ecology 8 (2), 153–61. o. DOI:10.1093/beheco/8.2.153.
- ↑ C.N. Spottiswoode, J.F.R. Colebrook-Robjent (2007). „Egg puncturing by the brood parasitic Greater Honeyguide and potential host counteradaptations”. Behavioral Ecology 18 (4), 792–799. o. DOI:10.1093/beheco/arm025.
- ↑ a b Edwards, DB (2012). „Immune investment is explained by sexual selection and pace-of-life, but not longevity in parrots (Psittaciformes).”. PLOS ONE 7 (12), e53066. o. DOI:10.1371/journal.pone.0053066. PMID 23300862. PMC 3531452.
- ↑ Doutrelant, C (2012. január 1.). „Female plumage coloration is sensitive to the cost of reproduction. An experiment in blue tits.”. Journal of Animal Ecology 81 (1), 87–96. o. DOI:10.1111/j.1365-2656.2011.01889.x. PMID 21819397.
- ↑ (2009) „The genetics of inbreeding depression”. Nat. Rev. Genet. 10 (11), 783–96. o. DOI:10.1038/nrg2664. PMID 19834483.
- ↑ (1987) „The molecular basis of the evolution of sex”. Adv. Genet. 24, 323–70. o. DOI:10.1016/s0065-2660(08)60012-7. PMID 3324702.
- ↑ Michod, R.E. (1994). "Eros and Evolution: A Natural Philosophy of Sex" Addison-Wesley Publishing Company, Reading, Massachusetts. ISBN 978-0201442328
- ↑ (2012) „Inbreeding causes early death in a passerine bird”. Nat Commun 3, 863. o. DOI:10.1038/ncomms1870. PMID 22643890.
- ↑ (2002) „Environmental conditions affect the magnitude of inbreeding depression in survival of Darwin's finches”. Evolution 56 (6), 1229–39. o. DOI:10.1111/j.0014-3820.2002.tb01434.x. PMID 12144022.
- ↑ S.A. Kingma, M.L. Hall, A. Peters (2013). „Breeding synchronization facilitates extrapair mating for inbreeding avoidance”. Behavioral Ecology 24 (6), 1390–1397. o. DOI:10.1093/beheco/art078.
- ↑ (2008) „Dispersal as a means of inbreeding avoidance in a wild bird population”. Proc. Biol. Sci. 275 (1635), 703–11. o. DOI:10.1098/rspb.2007.0989. PMID 18211876. PMC 2596843.
- ↑ (2012) „Inbreeding avoidance mechanisms: dispersal dynamics in cooperatively breeding southern pied babblers”. J Anim Ecol 81 (4), 876–83. o. DOI:10.1111/j.1365-2656.2012.01983.x. PMID 22471769.
- ↑ (2015) „How cooperatively breeding birds identify relatives and avoid incest: New insights into dispersal and kin recognition”. BioEssays 37 (12), 1303–8. o. DOI:10.1002/bies.201500120. PMID 26577076.
- ↑ a b M. Clout, J. Hay (1989). „The importance of birds as browsers, pollinators and seed dispersers in New Zealand forests”. New Zealand Journal of Ecology 12, 27–33. o.
- ↑ Gary Stiles, F. (1981). „Geographical Aspects of Bird-Flower Coevolution, with Particular Reference to Central America”. Annals of the Missouri Botanical Garden 68 (2), 323–51. o. DOI:10.2307/2398801. JSTOR 2398801.
- ↑ E. Temeles (2002). „The Role of Flower Width in Hummingbird Bill Length–Flower Length Relationships”. Biotropica 34 (1), 68–80. o. DOI:10.1111/j.1744-7429.2002.tb00243.x.
- ↑ S. Wainright, et al. (1998). „Utilization of nitrogen derived from seabird guano by terrestrial and marine plants at St. Paul, Pribilof Islands, Bering Sea, Alaska”. Marine Ecology 131 (1), 63–71. o. [2020. március 28-i dátummal az eredetiből archiválva]. DOI:10.1007/s002270050297. (Hozzáférés: 2017. március 29.)
- ↑ A. Bosman, A. Hockey (1986). „Seabird guano as a determinant of rocky intertidal community structure”. Marine Ecology Progress Series 32, 247–57. o. DOI:10.3354/meps032247.
- ↑ William J. Bond, William G. Lee, Joseph M. Craine (2004). „Plant structural defences against browsing birds: a legacy of New Zealand's extinct moas”. Oikos 104 (3), 500–08. o. DOI:10.1111/j.0030-1299.2004.12720.x.
- ↑ Bonney, Rick. Handbook of Bird Biology, Second, Princeton University Press (2004). ISBN 0-938027-62-X
- ↑ Dean W, Siegfried R, MacDonald I (1990). "The Fallacy, Fact, and Fate of Guiding Behavior in the Greater Honeyguide". Conservation Biology 4 (1) 99–101. Doi-PDF
- ↑ R. Singer, Y. Yom-Tov (1988). „The Breeding Biology of the House Sparrow Passer domesticus in Israel”. Ornis Scandinavica 19 (2), 139–44. o. DOI:10.2307/3676463. JSTOR 3676463.
- ↑ Dolbeer, R (1990). „Ornithology and integrated pest management: Red-winged blackbirds Agleaius phoeniceus and corn”. Ibis 132 (2), 309–22. o. DOI:10.1111/j.1474-919X.1990.tb01048.x.
- ↑ R. Dolbeer, J. Belant, J. Sillings (1993). „Shooting Gulls Reduces Strikes with Aircraft at John F. Kennedy International Airport”. Wildlife Society Bulletin 21, 442–50. o.
- ↑ "Will Wind Turbines Ever Be Safe for Birds?", by Emma Bryce, Audobon, US National Audobon Society, 16 March 2016. Accessed 19 March 2017.
- ↑ Reed, KD, et al. (2003). „Birds, Migration and Emerging Zoonoses: West Nile Virus, Lyme Disease, Influenza A and Enteropathogens”. Clinical medicine & research 1 (1), 5–12. o. DOI:10.3121/cmr.1.1.5. PMID 15931279. PMC 1069015.
- ↑ Brown, Lester. 3: Moving Up the Food Chain Efficiently., Outgrowing the Earth: The Food Security Challenge in an Age of Falling Water Tables and Rising Temperatures [archivált változat]. earthscan (2005). ISBN 978-1-84407-185-2. Hozzáférés ideje: 2017. március 30. [archiválás ideje: 2016. március 3.]
- ↑ A. Simeone, X. Navarro (2002). „Human exploitation of seabirds in coastal southern Chile during the mid-Holocene”. Rev. Chil. Hist. Nat 75 (2), 423–31. o. DOI:10.4067/S0716-078X2002000200012.
- ↑ Aidan Keane, M.de L. Brooke, P.J.K. McGowan (2005). „Correlates of extinction risk and hunting pressure in gamebirds (Galliformes)”. Biological Conservation 126 (2), 216–33. o. DOI:10.1016/j.biocon.2005.05.011.
- ↑ The Guano War of 1865–1866. World History at KMLA. (Hozzáférés: 2007. december 18.)
- ↑ R. Cooney. P. Jepson (2006). „The international wild bird trade: what's wrong with blanket bans?”. Oryx 40 (1), 18–23. o. DOI:10.1017/S0030605306000056.
- ↑ M. Manzi, O. T. Coomes (2002). „Cormorant fishing in Southwestern China: a Traditional Fishery under Siege. (Geographical Field Note)”. Geographic Review 92 (4), 597–603. o. DOI:10.2307/4140937. JSTOR 4140937.
- ↑ Pullis La Rouche, G. (2006). Birding in the United States: a demographic and economic analysis. Waterbirds around the world. Eds. G.C. Boere, C.A. Galbraith and D.A. Stroud. The Stationery Office, Edinburgh, UK. pp. 841–46. JNCC.gov.uk Archiválva 2011. március 4-i dátummal a Wayback Machine-ben, PDF
- ↑ Chamberlain, DE, et al. (2005). „Annual and seasonal trends in the use of garden feeders by birds in winter”. Ibis 147 (3), 563–75. o. DOI:10.1111/j.1474-919x.2005.00430.x. (Hozzáférés: 2017. március 31.)
- ↑ S. Routledge, K. Routledge (1917). „The Bird Cult of Easter Island”. Folklore 28 (4), 337–55. o. DOI:10.1080/0015587X.1917.9719006.
- ↑ Lukas, SE, et al. (1992). „Marihuana attenuates the rise in plasma ethanol levels in human subjects”. Neuropsychopharmacology 7 (1), 77–81. o. PMID 1326277.
- ↑ Ingersoll, Ernest (1923). Archive.org, "Birds in legend, fable and folklore". Longmans, Green and co. p. 214
- ↑ Thankappan Nair, P. (1974). „The Peacock Cult in Asia”. Asian Folklore Studies 33 (2), 93–170. o. DOI:10.2307/1177550. JSTOR 1177550.
- ↑ Smith, S. (2011). „Generative landscapes: the step mountain motif in Tiwanaku iconography.” (Automatic PDF download). Ancient America 12, 1–69. o. [2019. január 6-i dátummal az eredetiből archiválva]. (Hozzáférés: 2017. április 1.)
- ↑ Hauser, A. J. (1985). „Jonah: In Pursuit of the Dove”. Journal of Biblical Literature 104 (1), 21–37. o. DOI:10.2307/3260591. JSTOR 3260591.
- ↑ Meighan, C. W. (1966). „Prehistoric Rock Paintings in Baja California”. American Antiquity 31 (3), 372–92. o. DOI:10.2307/2694739. JSTOR 2694739.
- ↑ Tennyson A, Martinson P (2006). Extinct Birds of New Zealand Te Papa Press, Wellington ISBN 978-0-909010-21-8
- ↑ Clarke, CP (1908). „A Pedestal of the Platform of the Peacock Throne”. The Metropolitan Museum of Art Bulletin 3 (10), 182–83. o. DOI:10.2307/3252550. JSTOR 3252550.
- ↑ Boime, Albert (1999). „John James Audubon: a birdwatcher's fanciful flights”. Art History 22 (5), 728–55. o. DOI:10.1111/1467-8365.00184.
- ↑ Chandler, A. (1934). „The Nightingale in Greek and Latin Poetry”. The Classical Journal 30 (2), 78–84. o. JSTOR 3289944.
- ↑ Lasky, E. D. (1992. március 1.). „A Modern Day Albatross: The Valdez and Some of Life's Other Spills”. The English Journal 81 (3), 44–46. o. DOI:10.2307/820195. JSTOR 820195.
- ↑ Carson, A (1998). „Vulture Investors, Predators of the 90s: An Ethical Examination”. Journal of Business Ethics 17 (5), 543–55. o. DOI:10.1023/A:1017974505642.[halott link]
- ↑ Enriquez PL, Mikkola H (1997). "Comparative study of general public owl knowledge in Costa Rica, Central America and Malawi, Africa". pp. 160–66 In: J.R. Duncan, D.H. Johnson, T.H. Nicholls, (Eds). Biology and conservation of owls of the Northern Hemisphere. General Technical Report NC-190, USDA Forest Service, St. Paul, Minnesota. 635 pp.
- ↑ Lewis DP (2005). Owlpages.com, Owls in Mythology and Culture. Retrieved on 15 September 2007
- ↑ Dupree, N (1974). „An Interpretation of the Role of the Hoopoe in Afghan Folklore and Magic”. Folklore 85 (3), 173–93. o. DOI:10.1080/0015587X.1974.9716553. JSTOR 1260073.
- ↑ Matthew Head (1997). „Birdsong and the Origins of Music”. Journal of the Royal Musical Association 122 (1), 1–23. o. DOI:10.1093/jrma/122.1.1.
- ↑ Clark, Suzannah. Music Theory and Natural Order from the Renaissance to the Early Twentieth Century. Cambridge University Press (2001). ISBN 0-521-77191-9
- ↑ Reich, Ronni. „NJIT professor finds nothing cuckoo in serenading our feathered friends”, Star Ledger , 2010. október 15. (Hozzáférés: 2011. június 19.)
- ↑ Sibley, Charles. Phylogeny and classification of birds. New Haven: Yale University Press (1990). ISBN 0-300-04085-7
- ↑ Mayr, Ernst. Species Taxa of North American Birds: A Contribution to Comparative Systematics, Publications of the Nuttall Ornithological Club, no. 9. Cambridge, Mass.: Nuttall Ornithological Club (1970). OCLC 517185
- ↑ Jarvis, E.D., et al. (2014). „Whole-genome analyses resolve early branches in the tree of life of modern birds”. Science 346 (6215), 1320–1331. o. DOI:10.1126/science.1253451. PMID 25504713. PMC 4405904.
- ↑ Prum, R.O. et al. (2015) A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526, 569–573.
- ↑ BirdLife International announces more Critically Endangered birds than ever before. BirdLife International, 2009. május 14. [2013. június 17-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. május 15.)
- ↑ Kinver, Mark. „Birds at risk reach record high”, BBC News Online, 2009. május 13. (Hozzáférés: 2009. május 15.)
- ↑ Norris K, Pain D (eds, 2002). Conserving Bird Biodiversity: General Principles and their Application Cambridge University Press. ISBN 978-0-521-78949-3
- ↑ Brothers, NP (1991). „Albatross mortality and associated bait loss in the Japanese longline fishery in the southern ocean”. Biological Conservation 55 (3), 255–68. o. DOI:10.1016/0006-3207(91)90031-4.
- ↑ WD. urster, C. Wurster, W. Strickland (1965. július 1.). „Bird Mortality Following DDT Spray for Dutch Elm Disease”. Ecology 46 (4), 488–99. o. DOI:10.2307/1934880.; C. F. Wurster, D. H. Wurster, W. N. Strickland (1965). „Bird Mortality after Spraying for Dutch Elm Disease with DDT”. Science 148 (3666), 90–91. o. DOI:10.1126/science.148.3666.90. PMID 14258730.
- ↑ Blackburn, T, et al. (2004. szeptember 24.). „Avian Extinction and Mammalian Introductions on Oceanic Islands”. Science 305 (5692), 1955–58. o. DOI:10.1126/science.1101617. PMID 15448269.
- ↑ Fuller E (2000). Extinct Birds (2nd ed.). Oxford University Press, Oxford, New York. ISBN 0-19-850837-9
- ↑ Steadman D (2006). Extinction and Biogeography in Tropical Pacific Birds, University of Chicago Press. ISBN 978-0-226-77142-7
- ↑ S. Butchart, A. Stattersfield, N. Collar (2006). „How many bird extinctions have we prevented?”. Oryx 40 (3), 266–79. o. [2011. augusztus 10-i dátummal az eredetiből archiválva]. DOI:10.1017/S0030605306000950. (Hozzáférés: 2017. április 5.)
Fordítás
[szerkesztés]Ez a szócikk részben vagy egészben a Bird című angol Wikipédia-szócikk fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
További információk
[szerkesztés]- Magyar Madártani és Természetvédelmi Egyesület
- MME Kóka a Madarak Védelméért
- Magyarország madárfajai az iNaturaliston
- A világ madárfajai az iNaturaliston
- IUCN Vörös Lista
- Madárfotó galéria
- Kárpát-medencében megfigyelhető énekesmadarak
- Avionary – Western and Central Palaearctic Birds in 46 Languages
- Kongatják a vészharangot a tudósok, világszerte pusztulnak a ragadozó madarak (MTI/Hvg.hu, 2021-09-05)
Kapcsolódó szócikkek
[szerkesztés]- Madárnemek listája
- Magyarország madárfajainak listája
- A madarak hasznáról és káráról
- Madarak és fák napja
| Kezdetleges Archosauromorphák |  | |
|---|---|---|
| Crurotarsi archosaurusok | ||
| Avemetatarsalia és Ornithodira archosaurusok | Scleromochlus • Pteroszauruszok (Pterosauria) • Dinosauromorpha • Dinoszauruszok (Dinosauria) • Madármedencéjűek (Ornithischia) • Hüllőmedencéjűek (Saurischia) • Madarak (Aves) | |
| Repülő archosaurusok | Avialae • Archaeopteryx • Confuciusornis • Ichthyornis • Enantiornithes • Hesperornithes • Neornithes • Futómadár-szabásúak (Paleognathae) • Újmadárszabásúak (Neognathae) | |