Espermatocito
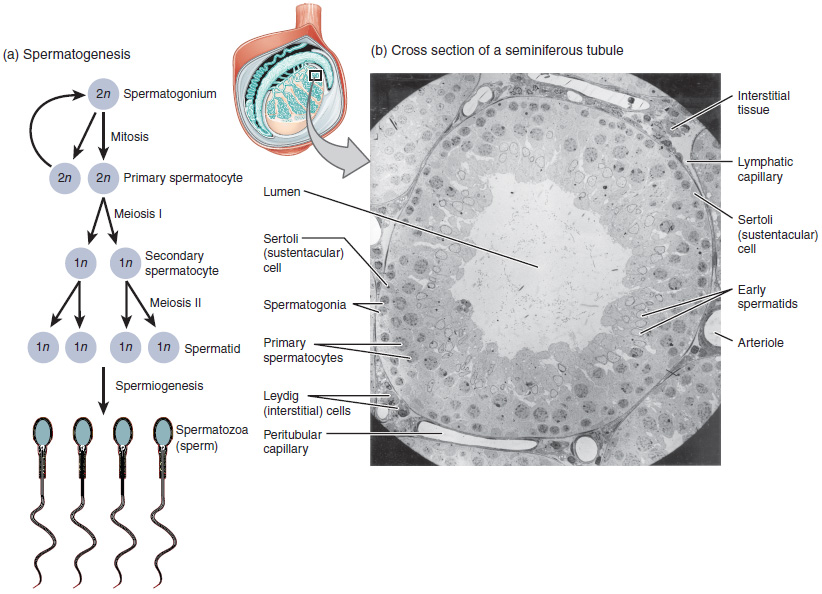
Os espermatocitos[1] son un tipo de gametocito masculino dos animais. Derivan de células xerminais inmaturas chamadas espermatogonias. Encóntranse nos testículos nunha estrutura chamada túbulos seminíferos.[2] Hai dous tipos de espermatocitos: primarios e secundarios. Os espermatocitos primarios son células diploides que se forman a partir de espermatogonias polo proceso mitótico da espermatocitoxénese. Os espermatocitos primarios sofren a primeira división meiótica dando lugar a espermatocitos secundarios haploides con cromosomas de dúas cromátides. Despois, os espermatocitos secundarios realizan a segunda división meiótica orixinando espermátides haploides con cromosomas dunha cromátide.[3][2]
En todos os animais os machos producen espermatocitos, incluso en animais hermafroditas como o verme nematodo Caenorhabditis elegans, que pode existir como macho ou como hermafrodita. Nos C. elegans hermafroditas prodúcese primeiro unha produción de espermatozoides, que é almacenada na súa espermateca. Unha vez que se forman os ovos, estes poden ser autofecundados con dito esperma e producir ata 350 fillos.[4]
Desenvolvemento
[editar | editar a fonte]


Na puberdade, as espermatogonias localizadas nas paredes dos túbulos seminíferos dos testículos actívanse e empezan a dividirse por mitose, formando dous tipos de células A que posúen un núcleo oval cun nucléolo unido á envoltura nuclear; un dos tipos é o escuro ou Ad (dark) e o outro o pálido ou Ap (pale). As células Ad son espermatogonias que permanecen no compartimento basal (rexión externa do túbulo); estas células son unha reserva de células nais espermatogoniais que xeralmente non realizan mitoses. As de tipo Ap son células nais espermatogoniais que están dividíndose activamente, que empezan a diferenciarse a espermatogonias de tipo B, as cales teñen núcleos arredondados e heterocromatina unida á envoltura nuclear e ao centro do nucléolo.[5] As células de tipo B trasládanse ao compartimento adluminal (cara á rexión interna do túbulo) e convértense en espermatocitos primarios; este proceso complétase en 16 días.[3][6]
Os espermatocitos primarios son células diploides, que dentro do compartimeno adluminal continúan dividíndose iniciando a primeira división meiótica na que se orixinan dúas células fillas, os espermatocitos secundarios haploides (con cromosomas de dúas cromátides), proceso que dura 24 días. Cada espermatocito secundario forma dúas espermátidas durante a segunda división meiótica. As espermátidas son tamén células haploides, pero con cromosomas dunha soa cromátide.[2] Durante as divisións mitóticas e meióticas de espermatogonias e espermatocitos, a citocinese (división do citoplasma) é incompleta e as células quedan unidas por pontes citoplasmáticas formando sincitios. As espermátidas finais son tamén sincitios e, unha vez que maduran a espermatozoides, estes despréndense deste sincitio e pasan á luz do túbulo seminífero.
Aínda que os espermatocitos en división son sensibles á radiación e ao cancro, as células nais espermatogoniais non. Por tanto, despois de terminada unha terapia de radiación ou quimoterapia, as células nais espermatogoniais poden reiniciar a espermatoxénese.[7]

Papel das hormonas
[editar | editar a fonte]A formación de espermatocitos primarios (proceso chamado espermatocitoxénese) empeza nos humanos cando un home chega á madureza sexual na puberdade, arredor dos 10 a 14 anos en humanos.[8] Esta formación iníciase por emisións de pulsos da hormona hormona liberadora da gonadotropina (GnRH) no hipotálamo, o cal orixina a secreción de hormona estimulante dos folículos (FSH) e de hormona luteinizante (LH) producidas pola glándula pituitaria anterior. A liberación de FSH nos testículos impulsa a espermatoxénese e leva ao desenvolvemento de células de Sertoli, que actúan como células nutricias onde as espermátidas van madurar despois da 2ª división meiótica. A LH promove a secreción por parte das células de Leydig de testosterona nos testículos, que pasa ao sangue, induce a espermatoxénese e axuda á formación das características sexuais secundarias ao inicio da puberdade. De aquí en adiante, a secreción de FSH e LH (inducindo a produción de testosterona) estimulará a espermatoxénese ata que o home morra.[9] O incremento das hormonas FSH e LH en machos non incrementa a taxa de espermatoxénese. Porén, coa idade, a taxa de produción diminúe, mesmo se a cantidade de hormona segregada é constante; isto débese ás maiores taxas de dexeneración das células xerminais durante a profase meiótica.[2]
Resumo dos tipos celulares
[editar | editar a fonte]Na seguinte táboa preséntase a ploidía, número de copias e contaxe de cromosomas/cromátides dunha soa célula, xeralmente antes da síntese de ADN e a división (en G1 se é aplicable). Os espermatocitos primarios están detidos despois da síntese de ADN e antes da división.[2][3]
| Célula | Tipo | Ploidía/Cromosomas en humanos | Número de copias de ADN/cromátides en humanos | Proceso no que entra a célula | Duración |
|---|---|---|---|---|---|
| espermatogonia (tipos Ad, Ap e B) | células xerminais | diploide (2N) / 46 | 2C / 46 | espermatocitoxénese (Mitose) | 16 días |
| espermatocito primario | gametocito masculino | diploide (2N) / 46 | 4C / 2x46 | espermatocitoxénese (Meiose I ) | 24 días |
| espermatocito secundario | gametocito masculino | haploide (N) / 23 | 2C / 46 | espermatidoxénese (Meiose II ) | Unhas poucas horas |
| espermátidas | gamétida masculna | haploide (N) / 23 | 1C / 23 | espermioxénese | 24 días |
| espermatozoides | esperma | haploide (N) / 23 | 1C / 23 | espermiación | 64 días (total) |
Fisioloxía
[editar | editar a fonte]Danos, reparación e insuficiencia
[editar | editar a fonte]Os espermatocitos sofren regularmente roturas de dobre febra no ADN e outros danos no ADN durante a profase da meiose. Estes danos poden orixinarse pola actividade programada do encima Spo11, un encima empregado na recombinación meiótica, así como por roturas non programadas no ADN, como as causadas por radicais libres oxidativos orixinados como produtos do metabolismo normal. Estes danos son reparados polas vías da recombinación homóloga e utilizan RAD1 e γH2AX, que recoñecen as roturas de dobre febra e modifican a cromatina, respectivamente. Como resultado, as roturas de dobre febra nas células meióticas, a diferenza de nas células mitóticas, conducen normalmente á apoptose ou morte celular.[10] A reparación por recombinación homóloga (HRR) de roturas de dobre febra ocorre en ratos durante as estapas secuenciais da espermatoxénese, pero é máis frecuente en espermatocitos.[11] Nos espermatocitos, os eventos HRR ocorren principalmente no paquiteno da profase meiótica e o tipo de HRR de conversión de xenes é o predominante, mentres que noutras etapas da espermatoxénese o tipo de HRR de intercambio recíproco é o máis frecuente.[11] Durante a espermatoxénese de rato, as frecuencias de mutación de células en diferentes etapas, incluíndo espaermatocitos en paquiteno, son de 5 a 10 veces menores que as frecuencias de mutación en células somáticas.[12] Debido á súa elevada capacidade de reparación do ADN, os espermatocitos probablemente xogan un papel central no mantemento destas taxas de mutación baixas e así na preservación da integridade xenética da liña xerminal masculina.
Os rearranxos cromosómicos heterocigotos causan unha alteración ou insuficiencia espermatoxénica; porén os mecanismos moleculares que o causan non se coñecen ben. Suxeriuse que unha posible causa é un mecanismo pasivo que implica un agrupamento de rexión asináptica nos espermatocitos. As rexións asinápticas están asociadas coa presenza de BRCA1, a quinase ATR e γH2AX nos espermatocitos en paquiteno.[13]
Mutacións específicas
[editar | editar a fonte]
O xene STRA8 (xene estimulado polo ácido retinoico 8) é necesario para o funcionamento da vía de sinalización do ácido retinoico en humanos, que conduce ao inicio da meiose. A expresión de STRA8 é maior en espermatocitos en preleptoteno (no estadio máis inicial da profase I da meiose) que en espermatogonias. Os espermatocitos mutantes en STRA8 poden iniciar a meiose, pero non poden completar o proceso. As mutacións en espermatocitos en leptoteno poden ter como resultado a condensación prematura dos cromosomas.[14]
As mutacións en Mtap2, unha proteína asociada aos microtúbulos, como a que se observa nos espermatocitos mutantes repro4, causa a detención do progreso da espermatoxénese durante a profase da 1ª división meiótica. Isto obsérvase por unha redución no número de espermátidas en mutantes repro4.[15]
As mutacións que afectan á recombinación poden ocorrer nos xenes Spo11, DMC1, ATM e MSH5 dos espermatocitos. Estas mutacións implican unha distorsión da reparación das roturas de dobre febra, que pode ter como resultado a detención da espermatoxénese na etapa IV do ciclo do epitelio seminífero.[16]
Historia
[editar | editar a fonte]
O proceso de espermatoxénese foi dilucidado ao longo de moitos anos cando os investigadores dividiron o proceso en múltiples etapas ou fases, e estudando a súa dependencia de factores intrínsecos (células xerminais e de Sertoli) e extrínsecos (hormonas FSH e LH).[17] O proceso da espermatoxénese en mamíferos no seu conxunto, que implica a transformación celular, mitose e meiose, foi ben estudado e documentado desde as décadas de 1950 a 1980. Porén, durante as décadas de 1990 e 2000 os investigadores centráronse en aumentar o coñecemento da regulación da espermatoxénese por xenes, proteínas e vías de sinalización, e nos mecanismos bioquímicos e moleculares que actúan no proceso. Máis recentemente, os efectos ambientais sobre a espermatoxénese convertéronse no foco de atención, a medida que a infertilidade masculina se fixo máis común.[18]
Un importante descubrimento no proceso de espermatoxénese foi a identificación do ciclo do epitelio seminífero en mamíferos, traballo realizado por C.P. Leblound e Y. Clermont en 1952, que estudaron as capas de espermatogonias, espermatocitos e espermátidas en túbulos seminíferos de ratas. Outro descubrimento crítico foi que a cadea hormonal hipotalámica-pituitaria-testicular desempeña un papel na regulación da espermatoxénese, o cal foi estudado por R. M. Sharpe en 1994.[18]
Outros animais
[editar | editar a fonte]
Os cilios primarios son orgánulos comúns nas células eucariotas e exercen un importante papel no desenvolvemento dos animais. Na mosca Drosophila os cilios primarios dos seus espermatocitos teñen propiedades únicas, xa que son ensamblados por catro centríolos independentemente na fase G2 e son sensibles a drogas que afectan os microtúbulos. Normalmente, os cilios primarios desenvólvense a partir dun centríolo na fase G0/G1 e non está afectados por drogas que afecten os microtúbulos.[19]
Mesostoma ehrenbergii é un platihelminto rabdocelo cunha distintiva etapa meiótica masculina durante a formación dos espermatocitos. No decurso da etapa de preanafase, aparecen sucos de estrangulamento nos espermatocitos en división que conteñen catro cromosomas univalentes. Ao final da anafase hai un en cada polo movéndose entre os polos do fuso sen que haxa interacción física entre eles (o que se chama distancia de segregación). Esta característica única permite aos investigadores estudar a forza que crean os polos do fuso, que fai que se movan os cromosomas, o control do suco de división e a distancia de segregación.[20][21]
Notas
[editar | editar a fonte]- ↑ Definición de espermatocito no Dicionario de Galego de Ir Indo e a Xunta de Galicia.
- ↑ 2,0 2,1 2,2 2,3 2,4 Boron, Walter F., MD, Ph.D., Editor; Boulpaep, Emile L. (2012). "54". Medical physiology a cellular and molecular approach (Print) (Updated second ed.). Philadelphia: Saunders Elsevier. ISBN 978-1-4377-1753-2.
- ↑ 3,0 3,1 3,2 Schöni-Affolter, Dubuis-Grieder, Strauch, Franzisk, Christine, Erik Strauch. "Spermatogenesis". Arquivado dende o orixinal o 01 de febreiro de 2022. Consultado o 22 de marzo de 2014.
- ↑ Riddle, DL; Blumenthal, T; Meyer, B.J.; et al., eds. (1997). "I, The Biological Model". C. elegans II (2nd ed.). Cold Spring Harbor. NY: Cold Spring Harbor Laboratory Press. Consultado o 13 de abril de 2014.
- ↑ Boitani, Carla; Di Persio, Sara; Esposito, Valentina; Vicini, Elena (2016-03-05). "Spermatogonial cells: mouse, monkey and man comparison". Seminars in Cell & Developmental Biology 59: 79–88. ISSN 1096-3634. PMID 26957475. doi:10.1016/j.semcdb.2016.03.002.
- ↑ Y, Clermont (1966). "Renewal of spermatogonia in man". American Journal of Anatomy 118 (2): 509–524. PMID 5917196. doi:10.1002/aja.1001180211.
- ↑ Tres, Abraham L. Kierszenbaum, Laura L. (2012). Histology and cell biology : an introduction to pathology (3ª ed.). Philadelphia, PA: Saunders. pp. Capítulo 20. ISBN 9780323078429.
- ↑ Starr, Taggart, Evers, Starr, Cecie, Ralph, Christine, Lisa (1 de xaneiro de 2012). Animal Structure & Function. Cengage Learning. p. 736. ISBN 9781133714071.
- ↑ Sherwood, Lauralee (2010). Human physiology : from cells to systems (7th ed.). Australia: Brooks/Cole, Cengage Learning. p. 751. ISBN 978-0495391845.
- ↑ Matulis S, Handel MA (agosto de 2006). "Spermatocyte responses in vitro to induced DNA damage". Molecular Reproduction and Development 73 (8): 1061–72. PMID 16700071. doi:10.1002/mrd.20508.
- ↑ 11,0 11,1 Srivastava N, Raman MJ (2007). "Homologous recombination-mediated double-strand break repair in mouse testicular extracts and comparison with different germ cell stages". Cell Biochem. Funct. 25 (1): 75–86. PMID 16989005. doi:10.1002/cbf.1375.
- ↑ Walter CA, Intano GW, McCarrey JR, McMahan CA, Walter RB (1998). "Mutation frequency declines during spermatogenesis in young mice but increases in old mice". Proc. Natl. Acad. Sci. U.S.A. 95 (17): 10015–9. Bibcode:1998PNAS...9510015W. PMC 21453. PMID 9707592. doi:10.1073/pnas.95.17.10015.
- ↑ Sciurano RB, Rahn MI, Rey-Valzacchi G, Coco R, Solari AJ (agosto de 2012). "The role of asynapsis in human spermatocyte failure". International Journal of Andrology 35 (4): 541–9. PMID 21977946. doi:10.1111/j.1365-2605.2011.01221.x.
- ↑ Mark, Manuel; Hugues Jacobs; Mustapha Oulad-Abdelghani; Christine Dennefeld; Betty Feret; Nadege Vernet; Carmen-Alina Codreanu; Pierre Chambon; Norbert Ghyselinck (7 de xullo de 2008). "STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo premature chromosome condensation". Journal of Cell Science 121 (19): 3233–3242. PMID 18799790. doi:10.1242/jcs.035071.
- ↑ Sun, Fengyun; Mary Ann Handel (10 de xaneiro de 2011). "A Mutation in Mtap2 is Associated with Arrest of Mammalian Spermatocytes before the First Meiotic Division". Genes 2 (1): 21–35. PMC 3909985. PMID 24501684. doi:10.3390/genes2010021.
- ↑ Barchi, Marco; S. Mahadevaiah; M. Di Giacomo; F. Baudat; D. de Rooij; P. Burgoyne; M. Jasin; S. Keeney (agosto de 2005). "Surveillance of Different Recombination Defects in Mouse Spermatocytes Yields Distinct Responses despite Elimination at an Identical Developmental Stage". Molecular and Cellular Biology 25 (16): 7203–7215. PMC 1190256. PMID 16055729. doi:10.1128/MCB.25.16.7203-7215.2005.
- ↑ Cheng, C. Yan, ed. (2008). Molecular mechanisms in spermatogenesis. New York: Springer Science+Business Media. pp. Chapter 1, page 1. ISBN 978-0-387-79990-2.
- ↑ 18,0 18,1 Cheng, C. Yan; Dolores D. Mruk (19 de abril de 2010). "The biology of spermatogenesis: the past, present and future". Phil. Trans. R. Soc. B. 1546 365 (1546): 1459–1463. PMC 2871927. PMID 20403863. doi:10.1098/rstb.2010.0024.
- ↑ Riparbelli MG, Cabrera OA, Callaini G, Megraw TL (2013). "Unique properties of Drosophila spermatocyte primary cilia". Biology Open 2 (11): 1137–47. PMC 3828760. PMID 24244850. doi:10.1242/bio.20135355.
- ↑ Ferraro-Gideon J, Hoang C, Forer A (xaneiro de 2014). "Meiosis-I in Mesostoma ehrenbergii spermatocytes includes distance segregation and inter-polar movements of univalents, and vigorous oscillations of bivalents". Protoplasma 251 (1): 127–43. PMID 23921676. doi:10.1007/s00709-013-0532-9.
- ↑ Ferraro-Gideon J, Hoang C, Forer A (setembro de 2013). "Mesostoma ehrenbergii spermatocytes--a unique and advantageous cell for studying meiosis". Cell Biology International 37 (9): 892–8. PMID 23686688. doi:10.1002/cbin.10130. hdl:10315/38106.
Véxase tamén
[editar | editar a fonte]Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]- Spermatogenesis Arquivado 18 de xuño de 2021 en Wayback Machine.
- The Male Reproductive System
- The Reproductive System Arquivado 17 de xuño de 2012 en Wayback Machine.