Arrestina

| |
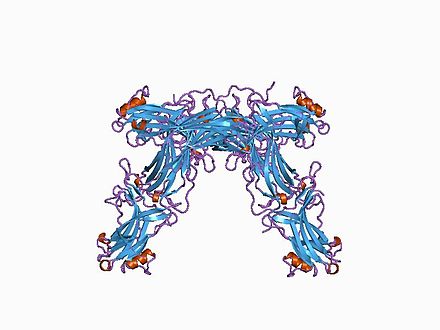
| Estrutura cristalográfica da arrestina S bovina.[1] | |
| Identificadores | |
| Símbolo | SAG |
| Símbolos alt. | arrestina-1 |
| Entrez | 6295 |
| HUGO | 10521 |
| OMIM | |
| RefSeq | NM_000541 |
| UniProt | P10523 |
| Outros datos | |
| Locus | Cr. 2 q37.1 |
| Identificadores | |
| Símbolo | ARRB1 |
| Símbolos alt. | ARR1, arrestina-2 |
| Entrez | 408 |
| HUGO | 711 |
| OMIM | |
| RefSeq | NM_004041 |
| UniProt | P49407 |
| Outros datos | |
| Locus | Cr. 11 q13 |
| Identificadores | |
| Símbolo | ? |
| Símbolos alt. | ARR2, arrestina-3 |
| Entrez | 409 |
| HUGO | 712 |
| OMIM | |
| RefSeq | NM_004313 |
| UniProt | P32121 |
| Outros datos | |
| Locus | Cr. 17 p13 |
| Identificadores | |
| Símbolo | ? |
| Símbolos alt. | ARRX, arrestina-4 |
| Entrez | 407 |
| HUGO | 710 |
| OMIM | |
| RefSeq | NM_004312 |
| UniProt | P36575 |
| Outros datos | |
| Locus | Cr. X q |
As arrestinas (abreviadas Arr) son unha pequena familia de proteínas importante na regulación da transdución de sinais en receptores acoplados á proteína G.[2][3] As arrestinas descubríronse como parte dun mecanismo de dous pasos conservado para regular a actividade dos receptores acoplados á proteína G (GPCRs) no sistema da rodopsina visual grazas aos traballos de Hermann Kühn, Scott Hall e Ursula Wilden[4] e no sistema β-adrenérxico en experimentos realizados por Martin J. Lohse e colaboradores.[5][6]
Función
[editar | editar a fonte]En resposta a un estímulo, os GPCRs activan a proteína G heterotrimérica. Para desactivar esta resposta ou adaptarse a un estímulo persistente, os receptores activos deben ser desensibilizados. O primeiro paso nesta desensibilización é a fosforilación do receptor por unha clase de serina/treonina quinases chamadas quinases do receptor acoplado á proteína G (GRKs). A fosforilación de GRK prepara especificamente o receptor activado para a unión da arrestina. Dita unión bloquea a sinalización mediada pola proteína G e etiqueta os receptores para a súa internalización e redirixe a sinalización a vías independentes da proteína G alternativas, como a sinalización da β-arrestina.[7][8][9][10][6] Ademais de aos GPCRs, as arrestinas únense a outras clases de receptores da superfice celular e a outras diversas proteínas de sinalización.[11]
Subtipos
[editar | editar a fonte]Os mamíferos expresan catro subtipos de arrestinas e cada un dos subtipos coñécese por múltiples nomes. O nome sistemático das arrestinas (do 1 ao 4) xunto cos alias máis amplamente usados de cada subtipo de arrestina indícanse en negras a continuación:
- Arrestina-1. Foi identificada orixinalmente co nome antíxeno S (SAG) que causaba uveíte (enfermidade autoinmune dos ollos); despois foi descrita independentemente como proteína de 48 kDa que se une á rodopsina fosforilada activada pola luz antes de que estivese claro que ambas as dúas eran a mesma proteína. Foi despois renomeada como arrestina visual, pero cando se clonou outro subtipo visual específico dos conos acuñouse para ela o termo arrestina dos bastóns. Este á súa vez resultou ser un nome incorrecto porque a arrestina-1 se expresa a niveis moi altos comparables tanto nas dúas células fotorreceptoras: bastóns e conos.
- Arrestina-2. Foi a primeira arrestina non visual clonada. Primeiro denominouse β-arrestina simplemente debido a que entre os dous GPCRs daquela dispoñibles en forma purificada, que eran a rodopsina e o receptor β2-adrenérxico, a proteína mostraba preferencia por este último.
- Arrestina-3. A segunda arrestina non visual clonada chamouse primeiro β-arrestina-2 (provocando retroactivamente o cambio do nome da β-arrestina a β-arrestina-1), incluso se nesa época estaba claro que as arrestinas non visuais interaccionaban con centos de GPCRs diferentes e non só co receptor β2-adrenérxico. Pouco despois, propuxéronse, respectivamente, os nomes sistemáticos arrestina-2 e arrestina-3.
- Arrestina-4. Clonárona dous grupos de investigadores e nomeárona arrestina dos conos, polo tipo de fotorreceptor no que se expresaba, e arrestina X (X-arrestin), polo cromosoma onde se encontraba o seu xene. Na base de datos HUGO o seu nome é arrestina-3.
Os peixes e outros vertebrados parece que teñen só tres arrestinas, xa que non se clonou ata agora ningún equivalente da arrestina-2, que é o subtipo máis abundante non visual nos mamíferos. O protocordado Ciona intestinalis (unha ascidia) ten só unha arrestina, que funciona como visual na súa larva móbil, que ten ollos moi desenvolvidos, e faise xenérica non visual no adulto sésil cego. As posicións conservadas de múltiples intróns do seu xene e os dos subtipos de arrestinas de vertebrados suxiren que todas evolucionaron a partir dunha arrestina ancestral.[12] Os invertebrados inferiores, como o nematodo Caenorhabditis elegans, tamén teñen unha soa arrestina. Os insectos teñen arr1 e arr2, denominadas orixinalmente “arrestinas visuais” porque se expresan en fotorreceptores, e un subtipo non visual (proteína kurtz en Drosophila[13]). Posteriormente descubríuse que arr1 e arr2 exercen un papel importante nas neuronas olfactorias e renomeáronse como “sensoriais”. Os fungos teñen parentes distantes das arrestinas implicados na percepción do pH.
Distribución nos tecidos
[editar | editar a fonte]Virtualmente en todas as células eucariotas prodúcese a expresión dunha ou máis arrestinas. En mamíferos, a arrestina-1 e a arrestina-4 están fundamentealmente confinadas nos fotorreceptores, mentres que a arrestina-2 e a arrestina-3 son ubicuas. As neuronas teñen o maior nivel de expresión de ambos os subtipos non visuais. En precursores neuronais exprésanse ambas a niveis comparables, mentres que en neuronas maduras a arrestina-2 está presente a niveis de 10 a 20 veces superiores aos da arrestina-3.
Mecanismo
[editar | editar a fonte]As arrestinas bloquean o acoplamento do GPCR ás proteínas G de dúas maneiras. Primeiro, a unión das arrestinas á cara citoplasmática do receptor agocha o sitio de unión para a proteína G heterotrimérica, impedindo a súa activación (desensibilización).[14] Segundo, a arrestina liga o receptor a elementos da maquinaria de internalización, a clatrina e o adaptador da clatrina AP2, que promoven a internalización do receptor por medio de depresións revestidas e o posterior transporte a compartimentos internos chamados endosomas. Seguidamente, o receptor pode ser dirixido a compartimentos de degradación (lisosomas) ou reciclado de volta á membrana plasmática, onde pode de novo sinalizar. A forza da interacción arrestina-receptor xoga un papel nesta elección: os complexos máis fortemente unidos adoitan incrementar a probabilidade da degradación do receptor (clase B), mentres que os complexos máis transitorios favorecen a reciclaxe (clase A), aínda que esta “regra” non é absoluta.[2] Máis recentemente as interaccións directas entre as proteínas G da familia Gi/o e a arrestina descubríronse augas abaixo de múltiples receptores, independentemente do acoplamento de proteínas G canónicas.[15] Estes descubrimentos recentes presentan un mecanismo de sinalización do GPCR distinto da activación das proteínas G canónicas e a desensibilización de β-arrestina, no cal os GPCRs causan a formación de complexos de sinalización Gαi:β-arrestina.
Estrutura
[editar | editar a fonte]As arrestinas son moléculas alongadas, nas cales varias interaccións intramoleculares manteñen a orientación relaiva dos seus dous dominios. As arrestinas de células non estimuladas están localizadas no citoplasma nunha conformación basal “inactiva”. Os GCPRs fosforilados activos recrutan a arrestina á membrana plasmática. A unión do receptor induce un cambio conformacional global que implica o movemento dos dous dominios da arrestina e a liberación da súa cola C-terminal que contén sitios de unión para a clatrina e AP2. O aumento da accesibilidade destes sitios en arrestinas unidas a receptor etiqueta o complexo arrestina-receptor á depresión revestida. As arrestinas tamén se unen aos microtúbulos (parte do citoesqueleto), onde adoptan aínda outra conformación, diferente das da forma libre e da forma unida a receptor. As arrestinas unidas a microtúbulos recrutan certas proteínas ao citoesqueleto, o que afecta a súa actividade e/ou rediríxea ás proteínas asociadas aos microtúbulos.
As arrestinas desprázanse entre o núcleo celular e o citoplasma. As súas funcións nucleares non se comprenden totalmente, pero demostrouse que os catro subtipos de arrestinas de mamíferos retiran do núcleo algunhas das proteínas ás que se unen, como a proteína quinase JNK3 ou a ubiquitina ligase Mdm2. As arrestinas tamén modifican a expresión xénica ao aumentaren a tanscrición de certos xenes.
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Notas
[editar | editar a fonte]- ↑ 1,0 1,1 PDB 1CF1; Hirsch JA, Schubert C, Gurevich VV, Sigler PB (abril de 1999). "The 2.8 A crystal structure of visual arrestin: a model for arrestin's regulation". Cell 97 (2): 257–69. PMID 10219246. doi:10.1016/S0092-8674(00)80735-7.
- ↑ 2,0 2,1 Moore CA, Milano SK, Benovic JL (2007). "Regulation of receptor trafficking by GRKs and arrestins". Annual Review of Physiology 69: 451–82. PMID 17037978. doi:10.1146/annurev.physiol.69.022405.154712.
- ↑ Lefkowitz RJ, Shenoy SK (abril de 2005). "Transduction of receptor signals by beta-arrestins". Science 308 (5721): 512–7. Bibcode:2005Sci...308..512L. PMID 15845844. doi:10.1126/science.1109237.
- ↑ Wilden U, Hall SW, Kühn H (marzo de 1986). "Phosphodiesterase activation by photoexcited rhodopsin is quenched when rhodopsin is phosphorylated and binds the intrinsic 48-kDa protein of rod outer segments". Proceedings of the National Academy of Sciences of the United States of America 83 (5): 1174–8. Bibcode:1986PNAS...83.1174W. PMC 323037. PMID 3006038. doi:10.1073/pnas.83.5.1174.
- ↑ Lohse MJ, Benovic JL, Codina J, Caron MG, Lefkowitz RJ (xuño de 1990). "beta-Arrestin: a protein that regulates beta-adrenergic receptor function". Science 248 (4962): 1547–50. Bibcode:1990Sci...248.1547L. PMID 2163110. doi:10.1126/science.2163110.
- ↑ 6,0 6,1 Gurevich VV, Gurevich EV (xuño de 2006). "The structural basis of arrestin-mediated regulation of G-protein-coupled receptors". Pharmacology & Therapeutics 110 (3): 465–502. PMC 2562282. PMID 16460808. doi:10.1016/j.pharmthera.2005.09.008.
- ↑ Smith JS, Lefkowitz RJ, Rajagopal S (xaneiro de 2018). "Biased signalling: from simple switches to allosteric microprocessors". Nature Reviews. Drug Discovery 17 (4): 243–260. PMC 5936084. PMID 29302067. doi:10.1038/nrd.2017.229.
- ↑ Cahill TJ, Thomsen AR, Tarrasch JT, Plouffe B, Nguyen AH, Yang F, et al. (febreiro de 2017). "Distinct conformations of GPCR-β-arrestin complexes mediate desensitization, signaling, and endocytosis". Proceedings of the National Academy of Sciences of the United States of America 114 (10): 2562–2567. Bibcode:2017PNAS..114.2562C. PMC 5347553. PMID 28223524. doi:10.1073/pnas.1701529114.
- ↑ Kumari P, Srivastava A, Banerjee R, Ghosh E, Gupta P, Ranjan R, Chen X, Gupta B, Gupta C, Jaiman D, Shukla AK (November 2016). "Functional competence of a partially engaged GPCR-β-arrestin complex". Nature Communications 7: 13416. Bibcode:2016NatCo...713416K. PMC 5105198. PMID 27827372. doi:10.1038/ncomms13416.
- ↑ Kumari P, Srivastava A, Ghosh E, Ranjan R, Dogra S, Yadav PN, Shukla AK (April 2017). "Core engagement with β-arrestin is dispensable for agonist-induced vasopressin receptor endocytosis and ERK activation". Molecular Biology of the Cell 28 (8): 1003–10. PMC 5391177. PMID 28228552. doi:10.1091/mbc.E16-12-0818.
- ↑ Gurevich VV, Gurevich EV (febreiro de 2004). "The molecular acrobatics of arrestin activation". Trends in Pharmacological Sciences 25 (2): 105–11. PMID 15102497. doi:10.1016/j.tips.2003.12.008.
- ↑ Gurevich EV, Gurevich VV (2006). "Arrestins: ubiquitous regulators of cellular signaling pathways". Genome Biology 7 (9): 236. PMC 1794542. PMID 17020596. doi:10.1186/gb-2006-7-9-236.
- ↑ Roman, G., He, J., & Davis, R. L. (2000). kurtz, a novel nonvisual arrestin, is an essential neural gene in Drosophila. Genetics, 155(3), 1281–1295. https://doi.org/10.1093/genetics/155.3.1281
- ↑ Kang Y, Zhou XE, Gao X, He Y, Liu W, Ishchenko A, et al. (xullo de 2015). "Crystal structure of rhodopsin bound to arrestin by femtosecond X-ray laser". Nature 523 (7562): 561–7. Bibcode:2015Natur.523..561K. PMC 4521999. PMID 26200343. doi:10.1038/nature14656.
- ↑ Smith JS, Pack TF, et al. (2021). "Noncanonical scaffolding of Gαi and β-arrestin by G protein–coupled receptors". Science 371 (Ahead of print): eaay1833. PMC 8005335. PMID 33479120. doi:10.1126/science.aay1833.
- ↑ Han M, Gurevich VV, Vishnivetskiy SA, Sigler PB, Schubert C (setembro de 2001). "Crystal structure of beta-arrestin at 1.9 A: possible mechanism of receptor binding and membrane Translocation". Structure 9 (9): 869–80. PMID 11566136. doi:10.1016/S0969-2126(01)00644-X.
Véxase tamén
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]Arrestin Medical Subject Headings (MeSH) na Biblioteca Nacional de Medicina dos EUA.