Gen
Un gen es una unidad de información[1] en un locus de ácido desoxirribonucleico (ADN) que codifica un producto génico, ya sea proteínas o ARN. Es la unidad molecular de la herencia genética,[2][3] pues almacena la información genética y permite transmitirla a la descendencia. Los genes se encuentran en los cromosomas, y cada uno ocupa en ellos una posición determinada llamada locus. El conjunto de genes de una especie se denomina genoma.
Para cada locus, pueden existir varios alelos posibles (es decir, pueden tener distintas formas con distintas consecuencias). Cuando los genes se encuentran muy próximos, es menos probable que se separen en el entrecruzamiento, es decir, no se segregan en forma independiente sino en bloque. Se denominan grupos de ligamiento al conjunto de genes situados en locus próximos que se transmiten en conjunto.
Molecularmente el gen es una secuencia de nucleótidos contiguos en la molécula de ADN (o de ARN en el caso de algunos virus) que contiene la información necesaria para la síntesis de una macromolécula con función celular específica, es decir, vinculados al desarrollo o funcionamiento de una función fisiológica. Generalmente estos productos son proteínas, previo paso por ARN mensajero (ARNm), pero también ARN no codificantes, como ARN ribosómico (ARNr), ARN de transferencia (ARNt) y muchos otros con funciones reguladoras o cuya función se va conociendo poco a poco.
Historia
[editar]

El concepto de gen ha ido variando a lo largo del tiempo, conforme ha avanzado la ciencia que lo estudia, la genética:
- Gregor Mendel en sus experimentos propuso la idea original del gen, aunque él no los denominó genes, sino factores, y vendrían a ser los responsables de la transmisión de los caracteres de una generación a la siguiente (lo que ahora llamamos genotipo). El gen mendeliano es una unidad de función, estructura, transmisión, mutación y evolución que se distribuye y ordena linealmente en los cromosomas.
- La palabra gen fue acuñada en 1909 por el botánico danés Wilhelm Johannsen[4] a partir de una palabra griega que significa "generar", refiriéndose a la unidad física y funcional de la herencia biológica.
- Hacia 1950, se impuso el concepto de gen como la cadena de ADN que dirige la síntesis de una proteína. Este es un concepto que proporciona una naturaleza molecular o estructural al gen. El gen codifica proteínas y debe tener una estructura definida por el orden lineal de sus tripletes o codones.
- Más tarde surge el concepto de gen como lo que actualmente se llama un cistrón:[5] la cadena de ADN capaz de dirigir la síntesis de un ARN que codifica para un polipéptido (Dogma central de la biología molecular). Este concepto surge al comprobar que la mayoría de las proteínas están formadas por más de una cadena polipeptídica y que cada una de ellas está codificada por un gen diferente.
- Actualmente se sabe que algunos genes codifican más de un polipéptido y que una proteína puede ser codificada por el conjunto de diferentes genes. La existencia de genes solapantes y el procesamiento alternativo rebaten la hipótesis de un gen → un polipéptido. Más bien debe proponerse la relación inversa, un polipéptido → un gen.[6] Además existen algunos genes que no codifican proteínas sino ARN con función propia (ARN transferentes y ARN ribosómicos, por ejemplo) y que no se traducen, por lo que no es necesaria la traducción para que un gen tenga una función determinada. El gen es, pues, la unidad mínima de función genética, que puede heredarse.
Concepto moderno del gen
[editar]A partir de la teoría original de Mendel de la determinación de caracteres físicos específicos (por.ej., el color de la flor) mediante partículas hereditarias discretas, el concepto de gen ha evolucionado gradualmente hacia el de unidad funcional. Esto fue anunciado por primera vez en 1945 por el genetista George Beadle (1903-1989), quien propuso que cada gen era específico: la hipótesis «un gen, una proteína». Fue modificada posteriormente cuando se comprendió que los genes podían determinar además proteínas no enzimáticas y también cadenas polipeptídicas individuales (subunidades proteicas) y los diversos tipos de ARN involucrados en la síntesis de proteínas. El desarrollo de nuevas técnicas en la década de los sesenta y ochenta, especialmente la secuenciación del ADN y la clonación de los genes, permitió a los genetistas moleculares desentrañar la estructura precisa de los genes hasta el nivel de las bases.
Tales técnicas aportan mucha información sobre cómo se activan y desactivan los genes y sobre otros aspectos de su expresión.
Tipos de genes
[editar]
Un gen es una secuencia o segmento de ADN necesario para la síntesis de ARN funcional, como el ARN de transferencia o el ARN ribosomal. Sin embargo, estos dos tipos de ARN no codifican proteínas, lo cual es hecho por el ARN mensajero. Para ello, la transcripción genera una molécula de ARN que posteriormente sufrirá traducción en los ribosomas, proceso por el cual se genera una proteína. Muchos genes se encuentran constituidos por regiones codificantes (exones) interrumpidas por regiones no codificantes (intrones) que son eliminadas en el procesamiento del ARN (splicing). En células procariotas esto no ocurre pues los genes de procariotas carecen de intrones. La secuencia de bases presente en el ARN determina la secuencia de aminoácidos de la proteína por medio del código genético.
Otros genes no son traducidos a proteína, sino que cumplen su función en forma de ARN. Entre estos, encontramos genes de ARN transferente, ARN ribosómico, ribozimas y otros ARN pequeños de funciones diversas.[8]
Algunos genes han sufrido procesos de mutación u otros fenómenos de reorganización y han dejado de ser funcionales, pero persisten en los genomas de los seres vivos. Al dejar de tener función, se denominan pseudogenes, que constituyen un recurso evolutivo para la especie, ya que son regiones de ADN quasifuncionales que pueden aceptar mutaciones (y generar nuevas funciones) sin perjuicio de las funciones que ya se desarrollan en el organismo, y pueden ser muy parecidos a otros genes del mismo organismo que sean funcionales.
Estructura y función
[editar]
Evolución molecular
[editar]Mutación
[editar]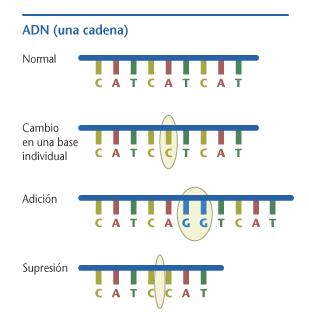
La replicación del ADN se hace con extrema precisión, sin embargo pueden producirse errores, llamados mutaciones.[3] La tasa de errores en las células eucariotas puede alcanzar tan solo una tasa de 10–8 mutaciones por nucleótido replicado,[9][10] mientras que en algunos virus de ARN la tasa puede subir hasta los 10−3.[11] Así, en cada generación cada genoma humano acumula aproximadamente entre 1 y 2 nuevas mutaciones.[11] Pequeñas mutaciones se pueden originar durante el proceso de replicación del ADN, que suelen repararse por un proceso conocido como reparación del ADN, aunque si este proceso no identificara o no corrigiera la mutación, se produciría un daño que nos llevaría a la aparición de mutaciones genéticas. Este fallo no es más que la sustitución de una base nitrogenada sustituida por otra, o bien algo mucho más peligroso, un desplazamiento del marco de lectura en las cuales se inserta o se elimina una base (o dos, el cambio afectaría menos si la deleción ocurre en tres bases, puesto que la lectura del ADN se hace por tripletes). Cualquiera de estas mutaciones pueden modificar la función del gen, dando lugar a una mutación sin sentido (cambios de un codón por otro que se traduce por un aminoácido diferente, que modifica la funcionalidad de la proteína traducida).[12] También puede originarse una mutación sin cambio de sentido, en la cual, el codón resultante se traduce en el mismo aminoácido (el código genético es degenerado, lo que viene a significar que varias combinaciones de tripletes se traducen en un mismo aminoácido): esto sería una mutación con sentido. Se pueden causar mutaciones más grandes por errores en la recombinación, en la sinapsis o durante el proceso de meiosis o mitosis durante el reparto cromosómico, causando mutaciones cromosómicas y aneuploidías, respectivamente. Estos fallos incluyen la duplicación génica (duplicación de la carga genética de una célula al contener una copia extra del cromosoma que lo alberga), o el borrado, la reorganización y la inversión de largas secciones de un cromosoma. También el mecanismo de reparación del ADN puede introducir errores cuando trata de reparar el daño físico a la molécula de ADN.[3]: 5.4
A la existencia de múltiples alelos de un gen, concurrentes en una misma población se le llama polimorfismo. La mayoría de alelos distintos funcionan de manera equivalente; sin embargo algunos puedan poseer caracteres biológicos distintos. El alelo más común de un gen se llama tipo salvaje, y a los alelos raros (infrecuentes) se les llama mutantes. La variación en frecuencias relativas de alelos diferentes en una población es consecuencia de la selección natural y la deriva genética (medidas del éxito adaptativo y del efecto estadístico respectivamente de un alelo).[13] El alelo del tipo salvaje no es definitivamente el antepasado de los alelos menos comunes ni es necesariamente cierto que estén más adaptados.
La mayoría de las mutaciones que ocurran en genes tienden a ser neutrales porque no afectan el fenotipo del organismo. Incluso si una mutación no es neutral puede conducir a una proteína funcionalmente idéntica, ya que la sustitución puede darse con un aminoácido equivalente o no perjudicial dentro de la estructura interna de la proteína que codifica. Sin embargo muchas mutaciones son perjudiciales cuando no claramente letales, siendo descartadas por la selección natural. Los trastornos genéticos resultan de mutaciones perniciosas y, a veces, debido a una mutación espontánea en el individuo afectado en una etapa temprana de su desarrollo, o bien heredados de uno o ambos progenitores en donde otra copia del gen asumía la función que el hijo hereda. Unas pocas benefician al organismo, mejorando la aptitud, e importan mucho porque ellas dirigen así la evolución adaptativa.[3]
Homología de secuencias
[editar]
Genes con un ancestro común más reciente, o sea unos abolengos evolutivos compartidos, se conocen como homólogos.[14] Estos genes aparezcan, o por la duplicación de genes adentro del genoma de un organismo, y se llaman genes paralogos, o resultan de divergencias de los genes después de un evento de especiación, se llaman genes ortologos,[3]: 7.6 y muchas veces desempeñen una función que es lo mismo o semejante en organismos relacionados. Se asume a menudo que las funciones de los genes ortologos se parezcan más que las de los genes paralogos, aunque las diferencias son muy pequeñas.[15][16]
Se puede medir la relación entre genes comparando su alineamiento de secuencias de ADN.[3]: 7.6 El grado de secuencia semejante entre genes homólogos se llama secuencia conservada. La mayoría de los cambios no afecten la función del gen y por esto los genes acumulan mutaciones con el paso del tiempo, por la evolución molecular neutralista. Además, cualquier selección en un gen hará que su secuencia diverja a un ritmo diferente. Los genes bajo selección estabilizadora están restringidos y, por lo tanto, cambian más lentamente, mientras que los genes bajo selección direccional cambian la secuencia más rápidamente. Las diferencias de secuencia entre genes pueden usarse para análisis filogenéticos para estudiar cómo evolucionaron esos genes y cómo se relacionan los organismos de los que provienen.[17][18]
Origen de nuevos genes
[editar]
La fuente más común de genes en las células eucariotas es la duplicación cromosómica, la cual crea variación en el número de copias de un gen que ya existe en el genoma.[19][20] Los genes resultantes (paralogos) luego quizás divergieran en su secuencia y también en su función. Grupos de genes formados de esta manera se llaman una familia génica. Las duplicaciones y perdidas genéticas adentro de una familia son comunes y representan una fuente mayor de la diversidad genética y la biodiversidad evolutiva.[21] A veces, la duplicación de genes dará como resultado una copia no funcional de un gen, o una copia que deba funcionar, pero por las mutaciones experimenta pérdidas de funciones; tales genes se llaman pseudogenes.[3]
Genomas
[editar]
El genoma es el total del material genético de un organismo e incluye los genes y también las secuencias no codificantes.[22]
Número de genes en algunas formas biológicas
[editar]El tamaño del genoma y el número de genes codificantes varían enormemente entre las formas biológicas. Los genomas más pequeños ocurren en los virus, los cuales pueden tener solo dos genes codificantes para codificar sus proteínas,[23] y viroides, los cuales actúan como un gen singular de ARN no codificante.[24] Por otra parte las plantas pueden tener los genomas muy grandes,[25] con arroz que contiene >46 000 genes codificantes de proteínas.[26] El número total de genes codificantes de proteínas (el proteoma de la Tierra) se estima como 5 millones de secuencias.[27]
Aunque el número de pares de bases de ADN en el genoma humano se conoce desde la década de los 1960, la estimación del número de genes se ha cambiado durante los años por razones como cambios en la definición de que es un gen, y mejoras en los métodos usados para detectar los genes. Predicciones teóricos iniciales del número de los genes humanos se alcanzaran dos millones.[28] Experimentos iniciales indicaron que fueron entre 50 000-100 000 transcripciones de genes (Marcador de secuencia expresadas).[29] Luego, las secuencias hechas en el Proyecto Genoma Humano indicaron que muchos de las transcripciones fueron variantes alternativas del mismo gen, y el número total de genes codificantes proteínas se bajó hasta –20 000[30] con 13 de los genes codificantes en el genoma de la mitocondria.[31] Del genoma humano solo 1-2 % consistan de genes codificantes de proteínas,[32] con los demás siendo ADN no codificante como intrones, retrotransposones, y ARN no codificante.[32][33]

| Forma biológica | N.º de genes | pares de bases |
|---|---|---|
| Plantas | <50000 | <1011 |
| Humanos | 20.000-25.000[34] | 3 × 109 |
| Moscas | 12000 | 1,6 × 108 |
| Hongos | 6000 | 1,3 × 107 |
| Bacterias | 500-6000 | 5 × 105 - 107 |
| Mycoplasma genitalium | 500 | 580.000 |
| Virus ADN | 10-300 | 5000-800 000 |
| Virus ARN | 1-25 | 1000-23 000 |
| Plásmidos | 30-100[35] | 3000-900 000 |
| Transposones | 1-10 | 2000-10 000 |
| Viroides | 0-1 | –500 |
| Priones | 0 | 0 |
Cambios en los genes
[editar]
Los organismos diploides disponen de dos juegos de cromosomas homólogos, cada uno de ellos proveniente de uno de los padres, cuyos gametos (creado por meiosis) se fusionaron hacia una célula conocido como un cigoto, durante la reproducción sexual.[36] Por ejemplo, los gametos (óvulo y espermatozoide) del ser humano solo contienen 23 cromosomas simples, (son haploides) pero ya terminado la división celular del cigoto creado por la fertilización en cuatro células, cada célula del nuevo bebé va a tener 23 pares de cromosomas, o sea 46 cromosomas, uno de los pares procedente de la madre y otra del padre.[37]
Algunas enfermedades como la anemia drepanocítica, se pueden ocasionar por un cambio en un solo gen. Los genes pueden aparecer en versiones diferentes, con pequeñas variaciones en su secuencia: es lo que se denomina alelos. Los alelos pueden ser dominantes o recesivos. Cuando una sola copia del alelo hace que se manifieste el rasgo o el fenotipo, el alelo es dominante. Cuando son precisas dos copias del alelo, para que se manifieste su efecto, el alelo es recesivo.
Regulación
[editar]Un gen es el conjunto de una secuencia determinada de nucleótidos de uno de los lados de la "escalera" del cromosoma referenciado. La secuencia puede llegar a formar proteínas, o serán inhibidas, dependiendo del programa asignado para la célula que aporte los cromosomas.[38]
Transferencia horizontal de genes
[editar]La transferencia horizontal de genes se refiere al movimiento de genes entre los seres vivos que no es a través de la transmisión vertical de (padres/madres a hijos). La transferencia horizontal de genes se da más comúnmente en los organismos unicelulares, pero también se da en los organismos pluricelulares.[39][40] Esta transferencia ha resultado ser un factor importante en la evolución de los seres vivos.[41] Aunque resulte sorpredente hablar de una transferencia de genes que no sea a través de la descendencia, a partir de los años 40 se acumularon evidencias de que los procariotas podían mover su material genético por medio de unas moléculas conocidas como plásmidos. Estas moléculas pueden transportar genes asociados con la fertilidad, fenotipo y metabolismo, inclusive pueden incorporar genes que ayudan a las bacterias a volverse resistentes a los antibióticos o genes virulentos que favorecen la patogenia de las bacterias. Los plásmidos no solo pueden transmitirse entre cepas procariotas sino también entre especies diferentes.[39][42]
Sin embargo los plásmidos no eran los únicos vectores de genes, ya que posteriormente se descubrió que los transposones secuencias de ADN capaces de moverse a diferentes partes del genoma podían transferirse entre organismos que viven en simbiosis como por ejemplo: los líquenes, invertebrados como los insectos o crustáceos, hongos, plantas, etc.[43] En los humanos se ha documentado casos de transferencia horizontal de genes con nematodos parasitarios por medio de transposones.[44] También se hizo evidente que los agentes virales (virus, viroides y virus satélite) durante su contagio e infección podían incorporar genes de un huésped y llevarlo al de otro huésped, incluyendo otras especies.[45] Los virus pueden considerarse el único medio de transferencia horizontal de genes más predominante entre los seres vivos aunque sean infectivos, además los virus desarrollan partículas proteicas (viriones) que le permiten fácilmente la movilidad entre sus huéspedes. También más recientemente se ha identificado que los virus pueden recibir genes adicionales de los virus satélite que dependen de ellos para la coinfección. A su vez los plásmidos y transposones pueden recibir genes adicionales de los integrones los cuales son casetes de genes con movilidad que se encuentran en los procariotas. Esto demuestra que el movimiento de genes no solo existe en los organismos celulares sino también entre las partículas vectoras. Todas estas moléculas o partículas se les ha denominado comúnmente el mobiloma o elementos genéticos móviles.[46]
Otros mecanismos de transferencia horizontal de genes incluyen la recombinación, la pérdida de genes, la duplicación y la creación de genes.[47] La transferencia horizontal de genes resulta ser una limitante para los evolucionistas dado que si dos especies han intercambiado genes a lo largo de la evolución pueden aparecer como iguales en un análisis filogenético sin tener que haber compartido un ancestro en común, porque ese gen es el mismo aunque muchos otros genes no sean similares. Por esta razón a veces es ideal usar otra información para inferir grandes filogenias como la presencia o ausencia de genes, o más común incluir un gran rango de genes para el análisis filogenético. Los análisis del proteoma también pueden ser útil para determinar las relaciones filogenéticas entre los organismos que sufren transferencias horizontales de genes o tienen alta tasa de mutación genética.[39][48]
Ingeniería genética
[editar]En la ingeniería genética se modifica el genoma de un organismo usando métodos biotecnológicos. Desde la década de los 1970, se han desarrollado técnicas que pueden agregar y editar genes específicos de organismos[49] Las técnicas de ingeniería de genoma más recientes utilizan enzimas nucleasas para fomentar la reparación de ADN específica en cromosomas, con tal de inactivar o editar un gen cuando el corte producido se repare.[50][51][52][53] Existe un concepto semejante, el cual es biología sintética y se suele referir al diseño genético extensivo de organismos.[54]
Hoy en día la ingeniería genética es una herramienta de rutina para la investigación de ciertos organismos modelo. Por ejemplo, se puede agregar genes a las bacterias fácilmente[55] mientras que linajes de ratón knockout con genes específicos inactivados son usados para investigar la función de esos genes.[56][57] Se han modificado muchos genes con fines relacionados con la agricultura, la medicina y la biotecnología industrial.
Para su aplicación en organismos multicelulares, por lo general se modifica genéticamente un embrión que luego crecerá hasta convertirse en un organismo genéticamente modificado adulto.[58] Por otra parte, el material genético de células de organismos adultos puede ser editado mediante técnicas de terapia génica para curar enfermedades con causas genéticas.
Véase también
[editar]Referencias
[editar]- ↑ Curtis, Helena; Schnek, Adriana (30 de junio de 2008). Curtis. Biología. Ed. Médica Panamericana. ISBN 9789500603348. Consultado el 19 de febrero de 2018.
- ↑ Slack, J.M.W. Genes-A Very Short Introduction. Oxford University Press 2014
- ↑ a b c d e f g Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002). Molecular Biology of the Cell (Fourth edición). Nueva York: Garland Science. ISBN 978-0-8153-3218-3.
- ↑ Baca, Laura Elena Lauría; Álvarez, Claudia Lorena Cantú (4 de agosto de 2015). Biología 2. Grupo Editorial Patria. ISBN 9786077442776. Consultado el 29 de noviembre de 2017.
- ↑ Rodríguez, Rosa María Ramos (1986). Crecimiento y proporcionalidad corporal en adolescentes mexicanas. UNAM. ISBN 9789688375242. Consultado el 29 de noviembre de 2017.
- ↑ Campbell, Neil A.; Reece, Jane B. (2007). Biología. Ed. Médica Panamericana. ISBN 9788479039981. Consultado el 29 de noviembre de 2017.
- ↑ Bolzer, Andreas; Kreth, Gregor; Solovei, Irina; Koehler, Daniela; Saracoglu, Kaan; Fauth, Christine; Müller, Stefan; Eils, Roland; Cremer, Christoph; Speicher, Michael R.; Cremer, Thomas (2005). «Three-Dimensional Maps of All Chromosomes in Human Male Fibroblast Nuclei and Prometaphase Rosettes». PLoS Biology 3 (5): e157. PMID 15839726. doi:10.1371/journal.pbio.0030157.
- ↑ Ruiz, José Sabán (4 de octubre de 2012). Control global del riesgo cardiometabólico: La disfunción endotelial como diana preferencial. Ediciones Díaz de Santos. ISBN 9788499692975. Consultado el 29 de noviembre de 2017.
- ↑ Nachman, M. W., Crowell, S. L. (septiembre de 2000). «Estimate of the mutation rate per nucleotide in humans». Genetics 156 (1): 297-304. PMC 1461236. PMID 10978293.
- ↑ Roach, J. C., Glusman, G., Smit, A. F., et al. (abril de 2010). «Analysis of genetic inheritance in a family quartet by whole-genome sequencing». Science 328 (5978): 636-9. PMC 3037280. PMID 20220176. doi:10.1126/science.1186802.
- ↑ a b Drake, J. W., Charlesworth, B., Charlesworth, D., Crow, J. F. (abril de 1998). «Rates of spontaneous mutation». Genetics 148 (4): 1667-86. PMC 1460098. PMID 9560386.
- ↑ «What kinds of gene mutations are possible?». Genetics Home Reference. United States National Library of Medicine. 11 de mayo de 2015. Consultado el 19 de mayo de 2015.
- ↑ Andrews, Christine A. (2010). «Natural Selection, Genetic Drift, and Gene Flow Do Not Act in Isolation in Natural Populations». Nature Education Knowledge. SciTable (Nature Publishing Group) 3 (10): 5.
- ↑ Patterson, C (noviembre de 1988). «Homology in classical and molecular biology.». Molecular biology and evolution 5 (6): 603-25. PMID 3065587.
- ↑ Studer, RA; Robinson-Rechavi, M (mayo de 2009). «How confident can we be that orthologs are similar, but paralogs differ?». Trends in genetics : TIG 25 (5): 210-6. PMID 19368988.
- ↑ Altenhoff, AM; Studer, RA; Robinson-Rechavi, M; Dessimoz, C (2012). «Resolving the ortholog conjecture: orthologs tend to be weakly, but significantly, more similar in function than paralogs.». PLoS computational biology 8 (5): e1002514. PMID 22615551.
- ↑ Emery, Laura. «Introduction to Phylogenetics». EMBL-EBI. Consultado el 19 de mayo de 2015.
- ↑ Mitchell, Matthew W.; Gonder, Mary Katherine (2013). «Primate Speciation: A Case Study of African Apes». Nature Education Knowledge. SciTable (Nature Publishing Group) 4 (2): 1.
- ↑ Guerzoni, D; McLysaght, A (noviembre de 2011). «De novo origins of human genes.». PLoS genetics 7 (11): e1002381. PMID 22102832.
- ↑ Reams, AB; Roth, JR (2 de febrero de 2015). «Mechanisms of gene duplication and amplification.». Cold Spring Harbor perspectives in biology 7 (2): a016592. PMID 25646380.
- ↑ Demuth, JP; De Bie, T; Stajich, JE; Cristianini, N; Hahn, MW (20 de diciembre de 2006). «The evolution of mammalian gene families.». PloS one 1: e85. PMID 17183716.
- ↑ Ridley, M. (2006). Genome. New York, NY: Harper Perennial. ISBN 0-06-019497-9
- ↑ Belyi, V. A.; Levine, A. J.; Skalka, A. M. (22 de septiembre de 2010). «Sequences from Ancestral Single-Stranded DNA Viruses in Vertebrate Genomes: the Parvoviridae and Circoviridae Are More than 40 to 50 Million Years Old». Journal of Virology 84 (23): 12458-12462. doi:10.1128/JVI.01789-10.
- ↑ Flores, Ricardo; Di Serio, Francesco; Hernández, Carmen (febrero de 1997). «Viroids: The Noncoding Genomes». Seminars in Virology 8 (1): 65-73. doi:10.1006/smvy.1997.0107.
- ↑ Zonneveld, B. J. M. (2010). «New Record Holders for Maximum Genome Size in Eudicots and Monocots». Journal of Botany 2010: 1-4. doi:10.1155/2010/527357.
- ↑ Yum, J., Hu, S., Wang, J., Wong, G. K., Li, S., Liu, B., Deng, Y., Dai, L., Zhou, Y., Zhang, X., Cao, M., Liu, J., Sun J., Tang J., Chen Y., Huang X., Lin W., Ye C., Tong W., Cong L, Geng J, Han Y, Li L, Li W, Hu G, Huang X, Li W, Li J, Liu Z, Li L, Liu J, Qi Q, Liu J, Li L, Li T., Wang X., Lu H., Wu T., Zhu M., Ni P., Han H., Dong W., Ren X., Feng, X., Cui, P., Li, X., Wang, H., Xu, X., Zhai, W., Xu, Z., Zhang, J., He, S., Zhang, J., Xu, J., Zhang, K., Zheng, X., Dong, J., Zeng, W., Tao, L., Ye, J., Tan, J., Ren, X., Chen, X., He, J., Liu, D., Tian, W., Tian, C., Xia, H., Bao, Q., Li, G., Gao, H., Cao, T., Wang, J., Zhao, W., Li, P., Chen, W., Wang, X., Zhang, Y., Hu, J., Wang, J., Liu, S., Yang, J., Zhang, G., Xiong, Y., Li, Z., Mao, L., Zhou, C., Zhu, Z., Chen, R., Hao, B., Zheng, W., Chen, S., Guo, W., Li, G., Liu, S., Tao, M., Wang, J., Zhu, L, Yuan, L., Yang, H. (April 2002). «A draft sequence of the rice genome (Oryza sativa L. ssp. indica)». Science 296 (5565): 79-92. Bibcode:2002Sci...296...79Y. PMID 11935017. doi:10.1126/science.1068037.
- ↑
- ↑ Kauffman, S. A. (1969). «Metabolic stability and epigenesis in randomly constructed genetic nets». Journal of Theoretical Biology (Elsevier) 22 (3): 437-467. PMID 5803332. doi:10.1016/0022-5193(69)90015-0.
- ↑ Schuler GD, Boguski MS, Stewart EA, Stein LD, Gyapay G, Rice K, White RE, Rodriguez-Tomé P, Aggarwal A, Bajorek E, Bentolila S, Birren BB, Butler A, Castle AB, Chiannilkulchai N, Chu A, Clee C, Cowles S, Day PJ, Dibling T, Drouot N, Dunham I, Duprat S, East C, Edwards C, Fan JB, Fang N, Fizames C, Garrett C, Green L, Hadley D, Harris M, Harrison P, Brady S, Hicks A, Holloway E, Hui L, Hussain S, Louis-Dit-Sully C, Ma J, MacGilvery A, Mader C, Maratukulam A, Matise TC, McKusick KB, Morissette J, Mungall A, Muselet D, Nusbaum HC, Page DC, Peck A, Perkins S, Piercy M, Qin F, Quackenbush J, Ranby S, Reif T, Rozen S, Sanders C, She X, Silva J, Slonim DK, Soderlund C, Sun WL, Tabar P, Thangarajah T, Vega-Czarny N, Vollrath D, Voyticky S, Wilmer T, Wu X, Adams MD, Auffray C, Walter NA, Brandon R, Dehejia A, Goodfellow PN, Houlgatte R, Hudson JR, Ide SE, Iorio KR, Lee WY, Seki N, Nagase T, Ishikawa K, Nomura N, Phillips C, Polymeropoulos MH, Sandusky M, Schmitt K, Berry R, Swanson K, Torres R, Venter JC, Sikela JM, Beckmann JS, Weissenbach J, Myers RM, Cox DR, James MR, Bentley D, Deloukas P, Lander ES, Hudson TJ (October 1996). «A gene map of the human genome». Science 274 (5287): 540-6. Bibcode:1996Sci...274..540S. PMID 8849440. doi:10.1126/science.274.5287.540.
- ↑ Pertea, Mihaela; Salzberg, Steven L (2010). «Between a chicken and a grape: estimating the number of human genes». Genome Biology 11 (5): 206. doi:10.1186/gb-2010-11-5-206.
- ↑ Anderson, S.; Bankier, A. T.; Barrell, B. G.; de Bruijn, M. H. L.; Coulson, A. R.; Drouin, J.; Eperon, I. C.; Nierlich, D. P.; Roe, B. A.; Sanger, F.; Schreier, P. H.; Smith, A. J. H.; Staden, R.; Young, I. G. (9 de abril de 1981). «Sequence and organization of the human mitochondrial genome». Nature 290 (5806): 457-465. doi:10.1038/290457a0.
- ↑ a b Claverie JM (September 2005). «Fewer genes, more noncoding RNA». Science 309 (5740): 1529-30. Bibcode:2005Sci...309.1529C. PMID 16141064. doi:10.1126/science.1116800.
- ↑ Carninci, P., Hayashizaki, Y. (April 2007). «Noncoding RNA transcription beyond annotated genes». Current Opinion in Genetics & Development 17 (2): 139-44. PMID 17317145. doi:10.1016/j.gde.2007.02.008.
- ↑ «Human Genome Project Information» (en inglés). Archivado desde el original el 2 de enero de 2013. Consultado el 4 de febrero de 2010.
- ↑ Plasmids. Science Direct.
- ↑ (COORD.), A. FIGUERAS HUERTA (COORD ), P. MARTÍNEZ PORTELA (2012). GENETICA Y GENOMICA EN ACUICULTURA TOMO I GENETICA. Editorial Paraninfo. ISBN 9788484766339. Consultado el 29 de noviembre de 2017.
- ↑ Rice, F. Philip (1997). Desarrollo humano: estudio del ciclo vital. Pearson Educación. ISBN 9789688808085. Consultado el 29 de noviembre de 2017.
- ↑ Investigación y ciencia. Temas 38: La nueva genética. Pág. 46. Los peligros del dogmatismo. Por W. Wayt Gibbs (adaptación para Wikipedia).
- ↑ a b c Gogarten, Peter (2000). «Horizontal Gene Transfer: A New Paradigm for Biology». Esalen Center for Theory and Research Conference. Archivado desde el original el 21 de julio de 2012. Consultado el 20 de septiembre de 2020.
- ↑ Lin Edwards (4 de octubre de 2010). «Horizontal gene transfer in microbes much more frequent than previously thought». PhysOrg.com. Consultado el 6 de enero de 2012.
- ↑ Gyles, C; Boerlin P (marzo de 2014). «Horizontally transferred genetic elements and their role in pathogenesis of bacterial disease». Veterinary Pathology 51 (2): 328-340. PMID 24318976. doi:10.1177/0300985813511131.
- ↑ Varga M, Kuntova L, Pantucek R, Maslanova I, Ruzickova V, Doskar J (2012). «Efficient transfer of antibiotic resistance plasmids by transduction within methicillin-resistant Staphylococcus aureus USA300 clone». FEMS Microbiol. Lett. 332 (2): 146-152. PMID 22553940. doi:10.1111/j.1574-6968.2012.02589.x.
- ↑ McDonald, Megan C.; Taranto, Adam P.; Hill, Erin; Schwessinger, Benjamin; Liu, Zhaohui; Simpfendorfer, Steven; Milgate, Andrew; Solomon, Peter S. (29 de octubre de 2019). «Transposon-Mediated Horizontal Transfer of the Host-Specific Virulence Protein ToxA between Three Fungal Wheat Pathogens». mBio 10 (5). ISSN 2150-7511. PMC 6737239. PMID 31506307. doi:10.1128/mBio.01515-19.
- ↑ Muñoz-López, Martín; García-Pérez, José L. (April 2010). «DNA Transposons: Nature and Applications in Genomics». Current Genomics 11 (2): 115-128. ISSN 1389-2029. PMC 2874221. PMID 20885819. doi:10.2174/138920210790886871.
- ↑ Victor J Freeman (1951). «Studies on the virulence of bacteriophage-infected strains of Corynebacterium Diphtheriae». Journal of Bacteriology 61 (6): 675-688. PMC 386063. PMID 14850426.
- ↑ Viruses and mobile elements as drivers of evolutionary transitions. NCBI.
- ↑ Jain, R.; Rivera, M. C.; Lake, J. A. (30 de marzo de 1999). «Horizontal gene transfer among genomes: the complexity hypothesis». Proceedings of the National Academy of Sciences of the United States of America 96 (7): 3801-3806. ISSN 0027-8424. PMID 10097118. doi:10.1073/pnas.96.7.3801.
- ↑ Graham Lawton Why Darwin was wrong about the tree of life New Scientist Magazine issue 2692 21 January 2009 Accessed February 2009
- ↑ Stanley N. Cohen and Annie C. Y. Chang (1 de mayo de 1973). «Recircularization and Autonomous Replication of a Sheared R-Factor DNA Segment in Escherichia coli Transformants — PNAS». Pnas.org. Consultado el 17 de julio de 2010.
- ↑ Esvelt, K. M.; Wang, H. H. (2013). «Genome-scale engineering for systems and synthetic biology». Mol Syst Biol 9 (1): 641. PMC 3564264. PMID 23340847. doi:10.1038/msb.2012.66.
- ↑ Tan, W. S.; Carlson, DF.; Walton, M. W.; Fahrenkrug, SC.; Hackett, P. B. (2012). «Precision editing of large animal genomes». Adv Genet. Advances in Genetics 80: 37-97. ISBN 9780124047426. PMC 3683964. PMID 23084873. doi:10.1016/B978-0-12-404742-6.00002-8.
- ↑ Puchta, H.; Fauser, F. (2013). «Gene targeting in plants: 25 years later». Int. J. Dev. Biol 57 (6-7-8): 629-637. doi:10.1387/ijdb.130194hp.
- ↑ Ran, F. A., Hsu, P. D., Wright, J., Agarwala, V., Scott, D. A., Zhang, F. (2013). «Genome engineering using the CRISPR-Cas9 system». Nat Protoc 8 (11): 2281-308. PMC 3969860. PMID 24157548. doi:10.1038/nprot.2013.143.
- ↑ Kittleson, Joshua (2012). «Successes and failures in modular genetic engineering». Current Opinion in Chemical Biology. doi:10.1016/j.cbpa.2012.06.009.
- ↑ Berg, P.; Mertz, J. E. (2010). «Personal Reflections on the Origins and Emergence of Recombinant DNA Technology». Genetics 184 (1): 9-17. PMC 2815933. PMID 20061565. doi:10.1534/genetics.109.112144.
- ↑ Austin, Christopher P.; Battey, James F.; Bradley, Allan; Bucan, Maja; Capecchi, Mario; Collins, Francis S.; Dove, William F.; Duyk, Geoffrey et al. (September 2004). «The Knockout Mouse Project». Nature Genetics 36 (9): 921-924. ISSN 1061-4036. PMC 2716027. PMID 15340423. doi:10.1038/ng0904-921.
- ↑ «A review of current large-scale mouse knockout efforts – Guan – 2010 – genesis – Wiley Online Library». doi.wiley.com.
- ↑ Deng C (2007). «In celebration of Dr. Mario R. Capecchi's Nobel Prize». International Journal of Biological Sciences 3 (7): 417-419. PMID 17998949. doi:10.7150/ijbs.3.417. Archivado desde el original el 20 de mayo de 2012. Consultado el 16 de julio de 2015.
Bibliografía adicional
[editar]- Nicolás Jouve de la Barreda. Explorando los genes. Del big-bang a la nueva Biología. Ediciones Encuentro. ISBN 978-84-7490-901-2.