Tau-protein
| microtubule associated protein tau | |||||||
|---|---|---|---|---|---|---|---|
| Identifikatorer | |||||||
| Aliaser | PHF-taumicrotubule-associated protein tauneurofibrillary tangle proteintau proteinsprotein phosphatase 1regulatory subunit 103paired helical filament-tauG protein beta1/gamma2 subunit-interacting factor 1MAPTtau protein | ||||||
| External IDs | GeneCards: [1] | ||||||
| RNA-udtryksmønster | |||||||
 | |||||||
| Flere henvisninger til udtrykdata | |||||||
| Ortologer | |||||||
| Art | Menneske | Mus | |||||
| Entrez |
|
| |||||
| Ensembl |
|
| |||||
| UniProt |
|
| |||||
| RefSeq (mRNA) |
|
| |||||
| RefSeq (protein) |
|
| |||||
| Location (UCSC) | n/a | n/a | |||||
| PubMed-søgning | n/a | n/a | |||||
| Wikidata | |||||||
| |||||||

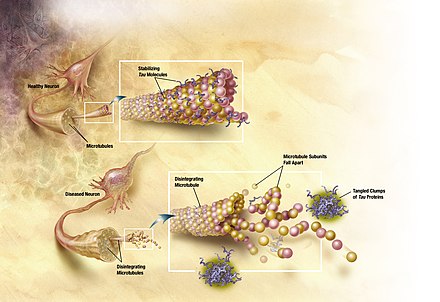

Tau-protein, Tau eller τ protein efter det græske bogstav Tau) er et protein, der stabiliserermikrotubuli i axoner og deltager i faciliteringen af exocytosen af synaptiske vesikler med neurotransmitter. Der er masser af tau i det centrale nervesystems neuroner, og der er færre andre steder, men der er også tau i meget små mængder i CNS's astrocyter og oligodendrocyter.[1] Tau findes som seks meget opløselige isoformer (varianter) med op til 441 aminosyrer. Isoformerne bliver produceret ved alternativ splejsning fra genet MAPT (mikrotubuli-associeret protein tau), som findes på kromosom 17.[2][3]
Tau-proteinets indre struktur er delvist uordnet, og neurodegenerativesygdomme som Alzheimers sygdom og Parkinsons sygdom,[4] er associeret med tau, der er blevet fejlfoldet, selvaggregeret og ikke længere stabiliserer mikrotubuli ordentligt, jvf. Proteinfejlfoldnings-sygdom eller proteinopati. Der kendes en del neurodegenerative sygdomme der kan føres tilbage til fejlfunktioner af tau, se her (en. Neurofibrillary tangle).
Funktion
[redigér | rediger kildetekst]En af taus hovedfunktioner er at modulere stabilitet af axonale mikrotubuli gennem isoformerne og fosforylering. Andre nervesystemers MAP'er kan udføre lignende funktioner. Det ses af mus uden tau, der ikke viste nogen abnormalitet i hjerneudviklingen - muligvis grundet en kompensering for manglende tau fra andre MAP'er.[5][6]
Struktur
[redigér | rediger kildetekst]Der findes seks tau-isoformer i det menneskelige hjernevæv, og de adskilles af deres antal af bindingsdomæner. Tre isoformer har tre bindingsdomæner, og de andre tre har fire bindingsdomæner. Bindingsdomænerne findes i proteinets carboxy-terminus, og de er positivt ladet (hvilket gør at de kan binde sig til de negativt ladede mikrotubuli). Isoformerne med fire bindingsdomæner er bedre til at stabilisere mikrotubuli, end dem med tre bindingsdomæner. Isoformerne er et resultat af alternativ deling i exonerne 2, 3 og 10 på tau-genet.
Tau er et fosforprotein med 79 potentielle serin (Ser) eller threonin (Thr) fosforyleringssteder på den største tau-isoform. Fosforylering rapporteres til at ske på cirka 30 af disse steder i normale tau-proteiner.[7] Forsforylering af tau reguleres af en masse kinaser, heriblandt PKN, en serin/theronin-kinase. Når PKN aktiveres, fosforyleres tau, hvilket resulterer i forstyrrelser i mikrotubuliorganiseringen.[8]
Fosforylering af tau reguleres også udviklingsmæssigt. Eksempelvis er foster-tau meget mere fosforyleret i det embryonale CNS end i det voksne tau.[9] Graden af fosforylering i alle seks isoformer nedsættes med alderen på grund af aktiveringen af fosfatase.[10] Lige som kinaser spiller fosfatase en rolle i reguleringen af fosforyleringen af tau. Eksempelvis er både PP2A og PP2B tilstede i menneskeligt hjernevæv og har evnen til at defosforylisere Ser396.[11] Bindingen af disse fosfataser til tau påvirker taus associering med MT'er.
Tau er desuden et intrinsikt uordnet protein uden en indre fast struktur, hvilket medfører stor fleksibilitet med mulighed for opbygning af store molekylære komplekser, der kan ses i mikroskopiske billeder af neurodegenererede hjerner (se billederne).
Genetik
[redigér | rediger kildetekst]Hos mennesker er MAPT-genet, der koder for tau-protein, lokaliseret på kromosom 17q21, indeholdende 16 exoner.[kilde mangler]. Den store tau-protein i hjernen er kodet af 11 exoner.[kilde mangler] Exon 2, 3 og 10 er alternativt delt, hvilket giver seks mulige kombinationer (2–3–10–; 2+3–10–; 2+3+10–; 2–3–10+; 2+3–10+; 2+3+10+). Derfor består tau-proteiner af en familie på seks isoformer men 352-441 aminosyrer, i den menneskelige hjerne. De adskiller sig fra enten nul, en eller to indførere af 29 aminosyrer ved N-terminus (exon 2 og 3), og tre eller fire gentagelses-regioner ved C-terminus-delen (exon 10). Så den længste isoform i centralnervesystemet har fire gentagelse (R1, R2, R3 og R4) og to indførere (441 aminosyrere i alt), mens den korteste isoform har tre gentagelse (R1, R3 og R4) og ingen indførere (352 aminosyrere i alt).
MAPT-genet har to haplogrupper, H1 og H2, i hvilke genet optræder med omvendt orientering. Haplogruppe H2 findes normalt i Europa og hos folk med europæisk baggrund. Haplogruppe H1 ser ud til at være associeret med øget sandsynlighed for bestemte demensformer, såsom Alzheimers sygdom. Tilstedeværelsen af begge haplogrupper i Europa betyder, at genkombinationen mellem omvendte haplotyper kan resultere i manglen på en af de fungerende kopier af genet, hvilket kan resultetere i medfødte defekter.[12][13][14][15]
Klinisk signifikans
[redigér | rediger kildetekst]Hyperfosforylering af tau-proteinet (tau inklusiner, pTau), kan resulterer i selvdannelse af knuder af parret heliske filamenter og lige filamenter, der er involveret i patogenesen ved Alzheimers sygdom, frontotemporal demens og andre tauopatier.[16]
Alle seks tau-isoformer er tilstede i et ofte hyperfosforyleret stadie i parrede heliske filamenter i hjerner hos Alzheimers-patienter. I andre neurodegenerative sygdomme, er samlede aflejringer beriget med bestemte tau-isoformer blev rapporteret. Når protein fejlfoldes kan det ellers meget opløselige protein danne ekstremt uopløselige produkter der er med til at give en del neurodegenerative sygdomme.
Undersøgelse har givet en idé om at tau måske udløses ekstracellulært af en eksosombaseret mekanisme i Alzheimers sygdom.[17][18]
Kønsspecifik tau-geneksprssion over forskellige regioner af den menneskelige hjerne har for nylig været impliceret i kønsforskelle i manifesteringen og risikoen for tauopatier.[19]
Nogle aspekter af hvordan sygdommen fungerer antyder også at det har ligheder med prion-proteiner.[20]
Traumatisk hjerneskade
[redigér | rediger kildetekst]Gentaget mild traumatisk hjerneskade (TBI), anderkendes nu som et centralt element i hjerneskade i kontaktsport, især amerikansk fodbold,[21][22] og den hjernerystelsesmæssige kræft ved militære eksplosioner,[23] kan føre til kronisk traumatisk encefalopati (CTE) der karakteriseres ved fibrillære knuder af hyperfosforyleret tau.[24]
Høje niveauer af tau protein i væsken hjernen ligger i, forbindes med dårlig helbredelse efter et hovedtraume.[25]
Alzheimers sygdoms tau-hypotese
[redigér | rediger kildetekst]Tau-hypotesen fortæller at store mængder eller abnormal fosforylering af tau resulterer i en transformation af normalt voksen tau til PHF-tau (parret helisk filament) og NFT'er (neurofibrillære knuder). Tau-protein er et stærk opløselig mikrotubuli-associeret protein (MAP).[6] Gennem dets isoformer og fosforylering interagerer tau-protein med tubulin for at stabilisere dannelsen af mikrotubuli. Tau-protein er en familie af seks isoformer, med mellem 352 og 441 aminosyrer. Den længste isoform i CNS har fire gentagelser (R1, R2, R3 og R4) og to indsættere (441 aminosyrere i alt), hvor den korteste isoform har tre gentagelser (R1, R3 og R4) og ingen indsættere (352 aminosyrer i alt). Alle seks tau-isoformer er tilstede i en ofte hyperfosforyleret stadie i parrede heliske filamenter fra AD.
Mutationer der ændrer funktionen og isoformudtrykket af tau fører til hyperfosforylering. Processen med samling af tau ved fraværet af mutationer er ikke kendt, men er muligvis et resultat af øget fosforylering, protease-aktivering eller eksponering for polyanioner, såsom glykoaminoglycans.[kilde mangler] Hyperfosforyleret tau ødelægger mikrotubuli og binder normalt tau, MAP 1 (mikrotubuliassocieret protein 1), MAP 2, og ubiquitin i knuder af PHF. Denne uopløselige struktur ødelægger cytoplastiske funktioner og forstyrre den aksonale transport, hvilket kan føre til celledød.[26]
Interaktioner
[redigér | rediger kildetekst]Tau-protein har vist sig at interagerer med proto-onkogene tyrosin-protein kinase:
Referencer
[redigér | rediger kildetekst]- ^ Shin RW, Iwaki T, Kitamoto T, Tateishi J (maj 1991). "Hydrated autoclave pretreatment enhances tau immunoreactivity in formalin-fixed normal and Alzheimer's disease brain tissues". Lab. Invest. 64 (5): 693-702. PMID 1903170.
- ^ Goedert M, Wischik CM, Crowther RA, Walker JE, Klug A (juni 1988). "Cloning and sequencing of the cDNA encoding a core protein of the paired helical filament of Alzheimer disease: identification as the microtubule-associated protein tau". Proc. Natl. Acad. Sci. U.S.A. 85 (11): 4051-5. doi:10.1073/pnas.85.11.4051. PMC 280359. PMID 3131773.
- ^ Goedert M, Spillantini MG, Jakes R, Rutherford D, Crowther RA (oktober 1989). "Multiple isoforms of human microtubule-associated protein tau: sequences and localization in neurofibrillary tangles of Alzheimer's disease". Neuron. 3 (4): 519-26. doi:10.1016/0896-6273(89)90210-9. PMID 2484340.
- ^ Lei P, Ayton S, Finkelstein DI, Adlard PA, Masters CL, Bush AI (november 2010). "Tau protein: relevance to Parkinson's disease". Int J Biochem Cell Biol. 42 (11): 1775-1778. doi:10.1016/j.biocel.2010.07.016. PMID 20678581.
- ^ Harada A, Oguchi K, Okabe S, Kuno J, Terada S, Ohshima T, Sato-Yoshitake R, Takei Y, Noda T, Hirokawa N (juni 1994). "Altered microtubule organization in small-calibre axons of mice lacking tau protein". Nature. 369 (6480): 488-91. doi:10.1038/369488a0. PMID 8202139.
- ^ a b Cleveland DW, Hwo SY, Kirschner MW (oktober 1977). "Purification of tau, a microtubule-associated protein that induces assembly of microtubules from purified tubulin". Journal of Molecular Biology. 116 (2): 207-25. doi:10.1016/0022-2836(77)90213-3. PMID 599557.
- ^ Billingsley ML, Kincaid RL (maj 1997). "Regulated phosphorylation and dephosphorylation of tau protein: effects on microtubule interaction, intracellular trafficking and neurodegeneration". Biochem. J. 323 (3): 577-91. PMC 1218358. PMID 9169588.
- ^ Taniguchi T, Kawamata T, Mukai H, Hasegawa H, Isagawa T, Yasuda M, Hashimoto T, Terashima A, Nakai M, Mori H, Ono Y, Tanaka C (marts 2001). "Phosphorylation of tau is regulated by PKN". J. Biol. Chem. 276 (13): 10025-31. doi:10.1074/jbc.M007427200. PMID 11104762.
- ^ Kanemaru K, Takio K, Miura R, Titani K, Ihara Y (maj 1992). "Fetal-type phosphorylation of the tau in paired helical filaments". J. Neurochem. 58 (5): 1667-75. doi:10.1111/j.1471-4159.1992.tb10039.x. PMID 1560225.
- ^ Mawal-Dewan M, Henley J, Van de Voorde A, Trojanowski JQ, Lee VM (december 1994). "The phosphorylation state of tau in the developing rat brain is regulated by phosphoprotein phosphatases". J. Biol. Chem. 269 (49): 30981-7. PMID 7983034.
- ^ Matsuo ES, Shin RW, Billingsley ML, Van deVoorde A, O'Connor M, Trojanowski JQ, Lee VM (oktober 1994). "Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau". Neuron. 13 (4): 989-1002. doi:10.1016/0896-6273(94)90264-X. PMID 7946342.
- ^ Shaw-Smith C, Pittman AM, Willatt L, Martin H, Rickman L, Gribble S, Curley R, Cumming S, Dunn C, Kalaitzopoulos D, Porter K, Prigmore E, Krepischi-Santos AC, Varela MC, Koiffmann CP, Lees AJ, Rosenberg C, Firth HV, de Silva R, Carter NP (september 2006). "Microdeletion encompassing MAPT at chromosome 17q21.3 is associated with developmental delay and learning disability". Nat. Genet. 38 (9): 1032-7. doi:10.1038/ng1858. PMID 16906163.
- ^ Zody MC, Jiang Z, Fung HC, Antonacci F, Hillier LW, Cardone MF, Graves TA, Kidd JM, Cheng Z, Abouelleil A, Chen L, Wallis J, Glasscock J, Wilson RK, Reily AD, Duckworth J, Ventura M, Hardy J, Warren WC, Eichler EE (september 2008). "Evolutionary toggling of the MAPT 17q21.31 inversion region". Nat. Genet. 40 (9): 1076-83. doi:10.1038/ng.193. PMC 2684794. PMID 19165922.
- ^ Almos PZ, Horváth S, Czibula A, Raskó I, Sipos B, Bihari P, Béres J, Juhász A, Janka Z, Kálmán J (november 2008). "H1 tau haplotype-related genomic variation at 17q21.3 as an Asian heritage of the European Gypsy population". Heredity (Edinb). 101 (5): 416-9. doi:10.1038/hdy.2008.70. PMID 18648385.
- ^ Hardy J, Pittman A, Myers A, Gwinn-Hardy K, Fung HC, de Silva R, Hutton M, Duckworth J (august 2005). "Evidence suggesting that Homo neanderthalensis contributed the H2 MAPT haplotype to Homo sapiens". Biochem. Soc. Trans. 33 (Pt 4): 582-5. doi:10.1042/BST0330582. PMID 16042549.
- ^ Alonso A, Zaidi T, Novak M, Grundke-Iqbal I, Iqbal K (juni 2001). "Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments". Proc. Natl. Acad. Sci. U.S.A. 98 (12): 6923-8. doi:10.1073/pnas.121119298. PMC 34454. PMID 11381127.
- ^ Hall, G.F. (2011) Tau misprocessing leads to non-classical tau secretion via vesicle release – implications for the spreading of tau lesions in AD Int Conf. Alz Dis. meeting Paris, France
- ^ Saman, S. and Hall, G. F. (2011) Tau secretion from M1C human neuroblastoma cells occurs via the release of exosomes. Keystone Meeting on Neurodegenerative diseases, Feb 2011, Taos NM
- ^ Koeglsberger S, Cordero-Maldonado ML, Antony P, Forster JI, Garcia P, Buttini M, Crawford A, Glaab E (november 2016). "Gender-Specific Expression of Ubiquitin-Specific Peptidase 9 Modulates Tau Expression and Phosphorylation: Possible Implications for Tauopathies". Molecular Neurobiology. in press: 1-15. doi:10.1007/s12035-016-0299-z. PMID 27878758.
- ^ Hall GF, Patuto BA (juli 2012). "Is tau ready for admission to the prion club?". Prion. 6 (3): 223-33. doi:10.4161/pri.19912. PMC 3399531. PMID 22561167.
- ^ "Brain Trauma". NOVA. PBS Online by WGBH.
- ^ Omalu, Bennet I. (1. juli 2005). "Chronic Traumatic Encephalopathy in a National Football League Player". Neurosurgery. 57: 128-134. doi:10.1227/01.NEU.0000163407.92769.ED. PMID 15987548. Arkiveret fra originalen 26. april 2017. Hentet 29. maj 2017.
- ^ Goldstein LE, Fisher AM, Tagge CA, Zhang XL, Velisek L, Sullivan JA, et al. (maj 2012). "Chronic traumatic encephalopathy in blast-exposed military veterans and a blast neurotrauma mouse model". Science Translational Medicine. 4 (134): 134ra60. doi:10.1126/scitranslmed.3003716. PMC 3739428. PMID 22593173.
- ^ McKee AC, Stern RA, Nowinski CJ, Stein TD, Alvarez VE, Daneshvar DH, et al. (januar 2013). "The spectrum of disease in chronic traumatic encephalopathy". Brain. 136 (Pt 1): 43-64. doi:10.1093/brain/aws307. PMC 3624697. PMID 23208308.
- ^ Magnoni S, Esparza TJ, Conte V, Carbonara M, Carrabba G, Holtzman DM, Zipfel GJ, Stocchetti N, Brody DL (april 2012) [first published online November 24, 2011]. "Tau elevations in the brain extracellular space correlate with reduced amyloid-β levels and predict adverse clinical outcomes after severe traumatic brain injury". Brain. 135 (Pt 4): 1268-80. doi:10.1093/brain/awr286. PMC 3326246. PMID 22116192. Lay summary – Washington University in St. Louis.
{{cite journal}}: Skabelonen anvender en forældet parameter|lay-url=(hjælp) - ^ Mudher M, Lovestone S (2002). "Alzheimer's disease- do tauists and Baptists finally shake hands?". Trends Neuroscience. 25: 22-6. doi:10.1016/s0166-2236(00)02031-2. PMID 11801334.
- ^ Jensen PH, Hager H, Nielsen MS, Hojrup P, Gliemann J, Jakes R (september 1999). "alpha-synuclein binds to Tau and stimulates the protein kinase A-catalyzed tau phosphorylation of serine residues 262 and 356". J. Biol. Chem. 274 (36): 25481-9. doi:10.1074/jbc.274.36.25481. PMID 10464279.
- ^ Giasson BI, Lee VM, Trojanowski JQ (2003). "Interactions of amyloidogenic proteins". Neuromolecular Med. 4 (1-2): 49-58. doi:10.1385/NMM:4:1-2:49. PMID 14528052.
- ^ Klein C, Kramer EM, Cardine AM, Schraven B, Brandt R, Trotter J (februar 2002). "Process outgrowth of oligodendrocytes is promoted by interaction of fyn kinase with the cytoskeletal protein tau". J. Neurosci. 22 (3): 698-707. PMID 11826099.
- ^ Yu WH, Fraser PE (april 2001). "S100beta interaction with tau is promoted by zinc and inhibited by hyperphosphorylation in Alzheimer's disease". J. Neurosci. 21 (7): 2240-6. PMID 11264299.
- ^ Baudier J, Cole RD (april 1988). "Interactions between the microtubule-associated tau proteins and S100b regulate tau phosphorylation by the Ca2+/calmodulin-dependent protein kinase II". J. Biol. Chem. 263 (12): 5876-83. PMID 2833519.
- ^ Hashiguchi M, Sobue K, Paudel HK (august 2000). "14-3-3zeta is an effector of tau protein phosphorylation". J. Biol. Chem. 275 (33): 25247-54. doi:10.1074/jbc.M003738200. PMID 10840038.