T-ćelija
| T-ćelia (Lymphocytus T) | |
|---|---|
 T-ćelija pod skenirajućim elektronskim mikroskopom | |
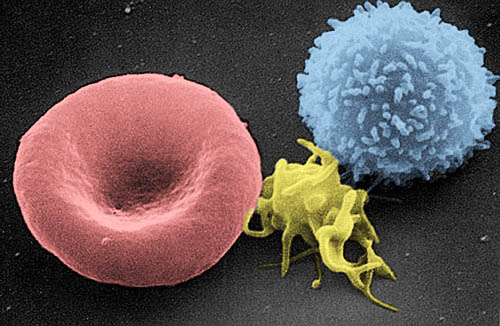 T-limfocit (desno), trombocit (centar) i crvena krvna ćelija (lijevo) | |
| Detalji | |
| Sistem | Imunski sistem |
| Identifikatori | |
| MeSH | D013601 |
| TH | TH {{{2}}}.html HH2.00.04.1.02007 .{{{2}}}.{{{3}}} |
| FMA | 62870 |
| Anatomska terminologija | |

T-ćelije ili T-limfociti su tip limfocita (podtipa bijelih krvnih zrnaca) koji imaju centralnu ulogu u ćelijskom imunitetu. Od drugih limfocita, kao što su B-ćelije i prirodnih ćelija ubica, razlikuju se po prisustvu receptorna T-ćelija na površini ćelije. One se nazivaju T-ćelije, jer sazrijevaju u timusnim timocitima[1] (neki također sazrijavaju krajnicima.[2]).
Od nekoliko podskupova T-ćelija, svaki ima različite funkcije. Većina ljudskih T-ćelija koje preuređuju svoje alfa i beta lance na ćelijskom receptoru, nazivaju se alfa i beta T-ćelije (αβ T-ćelije) i dio su adaptivnog imuniteta. Specijalizirane gama delta ćelije, (mali broj T-ćelija u ljudskom tijelu, mnogo češće kod preživara), imaju nevarirajuće receptore T-limfocita, s ograničenom raznolikosti, koji se mogu efektivno predstavljati antigene drugim T-ćelijama.[3] i smatraju se dijelom urođenog imunskog sistema.
Tipovi
[uredi | uredi izvor]Ćelije pomoćnice - Helperi
[uredi | uredi izvor]Helper T-ćelije (THćelije) – ćelije pomoćnici – pomažu ostalim bijelim krvnim ćelijama u imunskim procesima, uključujući sazrijevanje B ćelija u plazma ćelijama i memorijskim B ćelijama, te aktivaciji citotoksičnih T-ćelija i makrofaga. Ove ćelije su također poznate kao CD4+ T-ćelije, zbog ispoljavanja CD4 glikoproteina na površini. Helper T-ćelije postaju aktivne kada se prezentiraju peptidnim antigenima molekula MHC II klase, koje se ispoljavaju na površini antigen-prezentirajućih ćelija (APC). Jednom aktivirane, brzo se dijele i lučw male proteine zvane citokini, koji reguliraju ili pomažu aktivni imunski odgovor. Ove ćelije se mogu diferencirati u jedan od nekoliko podtipova, uključujući TH1, TH2, TH3, TH17, TH9, or TFH, koje luče različite citokine imunskog odgovora. Signalizacija iz APC usmjerava T-ćelije u posebne podstipove.
Citotoksične T-ćelije
[uredi | uredi izvor]Citotoksične T-ćelije (TC ćelije ili CTL) uništavaju ćelije koje su inficirane virusima i tumorskim ćelijamaan, a također su uključene i u reakcije na transplantacije organa. Ove ćelije su također poznate i kao CD8+ T-ćelije jer na površini ispoljavaju CD8 glikoprotein . Ove ćelije prepoznaju svoje ciljeve putem vezanja za antigen koji je asociran sa molekulama MHC klase I, koje se ispoljavaju na površini nukleiranih ćelija. Tokom sekrecije IL-10, adenozina i ostalih molekula koje se luče putem regulacijskih T-ćelija, CD8+ ćelije mogu preći u neaktivno stanje koje prevenira autoimunske bolesti.
Memorijske T-ćelije
[uredi | uredi izvor]Memorijske T-ćelije su podskup antigen – specifičnih T-ćelija koje traju dugoročno nakon savladavanja infekcije. One se brzo proširuju na veliki broj efektorskih T-ćelija na ponovno ispoljavanje svog srodnog antigena, omogućavajući da imunski "memorira" aktivnost protiv prošlih infekcija.[4] Memorijske T-ćelije obuhvataju tri podtipa:
- centralne memorijske T-ćelije(TCM ćelije),
- efektorske memoriske T-ćelije (TEM ćelije) i TEMRA ćelije,[5] te
- rezidentne memoriske T-ćelije (TRM).[6]
Supresorske ćelije
[uredi | uredi izvor]Supresorske T-ćelije su ključne za održavanje imunske tolerancije. Njihova glavna uloga je da ugase imunost koja je posredovana T-ćelijama, krajem imunske reakcije i da suzbiju autoreaktivne T-ćelije koje su izbjele proces negativne selekcije u timusu.
Supresorske T-ćelije, uz helper ćelije, po regulacijskoj funkciji, mogu se zajednički nazvati regulatorskim T-ćelijama.
Opisane su dvije glavne klase CD4+ Tregćelija cells have been described — FOXP3+ Treg i and FOXP3− Treg ćelije. Regulatorne T-ćelije mogu nastati u bilo kom dijelu toka normalnog razvoja u timusu, kada su poznate kao timusne Treg ćelije, ili mogu biti izazvane periferno i nazivaju se periferno izvedene Treg ćelije. Ova dva podskupa su ranije bila pod nazivom "prirodn" i "adaptivni" ili "inducirani".[7] Obje podskupine odgovaraju ekspresiji transkripcijskog faktora FOXP3 koji se može upotrebiti za identifikaciju ćelija mutacije gena za bolest FOXP3, koja se može prevenirati razvojem regulatornih T-ćelija, a koja je uzrok fatalne autoimunske bolesti IPEX.
Nekoliko drugih vrsta T-ćelija imaju supresivnu aktivnost, ali ne izražavaju FOXP3. To uključuje Tr1 i Th3 ćelije, za koje se smatra da nastaju tokom imunskog odgovora i aktom proizvodnje supresivnih molekula. Tr1 ćelije su povezane sa IL-10, Th3 ćelije su povezani sa TGF-beta. Nedavno, na ovu listu su dodate i Treg17 ćelije.[8]
Prirodne ćelije ubice
[uredi | uredi izvor]Prirodne T-ćelije ubice (NKT-ćelije - ne treba miješati sa prirodnim ćelijama ubicama urođenog imunskog sistema) premostiti adaptivni imuni sistem sa urođeni imuni sistem. Za razliku od konvencijskih T-ćelija, koje prepoznaju peptidne antigene prezentirne molekulama glavnim kompleksom histokompatibilnosti (MHC), NKT-ćelije prepoznaju glikolipidne antigen predstavljene molekulama pod nazivom CD1d. Kada se aktiviraju, ove ćelije mogu obavljati funkcije koje se pripisuju i Th i TC ćelijama (tj. proizvodnju citokina i oslobađanje citolitskih ćelija koje ubijaju molekule). Također su u stanju prepoznati i ukloniti neke tumorske ćelije i ćelije zaražene herpes virusima.
Nevarijantne sluzokožne ćelije
[uredi | uredi izvor]Sluzokožno povezani invarijantni T-limfociti (MAIT) su posebna vrsta T-ćelija, priznatih od oko 2009.[9][10][11][12]
Gama delta T-ćelije
[uredi | uredi izvor]Gama Delta T-ćelije (γδ T-limfociti) predstavljaju mali podskup T sćelija koje na površini imaju poseban receptor T-ćelije (TCR). Većina T-ćelija imaj receptor T-ćelija koji se sastoji od dva glikoproteinska lanca koji se zovu α- i β-TCR lanci. Međutim, u γδ T-limfocitima, TCR se sastoji od po jednog γ-lanca i δ-lanca. Ova grupa T-ćelija je manje uobičajena kod ljudi i miševa (oko 2% od ukupnog broja T-limfocita). Nalaze se uglavnom u sluznicama crijeva, unutar populacije limfocita poznate kao unutarepitelni limfociti. U kunića, ovaca i kokoši, broj γδ T-ćelija može biti visok i oko 60% od ukupnog broja T-limfocita. Antigene molekule koje aktiviraju γδ T-ćelije su još uvijek široko poznate. Međutim, γδ T-ćelije nisu ograničene MHCom i čini se da su u stanju da prepoznati cijeli protein, a ne zahtijevaju peptide koji će biti predstavljeni iz MHC molekula na transporteru. Neke Murinae (miševi), γδ T-ćelije prepoznaju MHC klase IB molekula. Ljudske Vγ9 / Vδ2 T-ćelije, koje čine najveći broj γδ T-limfocita u perifernoj krvi, su jedinstvene po tome što konkretno i brzo odgovaraju na niz nepeptidnih fosforiliziranih izoprenoidnik prekursora, skupnog naziva fosfoantigeni, koji se proizvode u gotovo svim živim ćelijama. Najčešći fosfoantigeni životinjskih i ljudskih ćelija (uključujući i ćelije raka) su izopentenil pirofosfati (IPP) i njegov izomer dimetilallil pirofosfat (DSPP). Mnogi mikrobi proizvode vrlo aktivni spoj hidroksi-DMAPP (HMB-PP) i odgovarajuće mononukleotidne konjugate, pored IPP i DMAPP. Biljne ćelije proizvode oba tipa fosfoantigena. Ljudske Vγ9/Vδ2 T-ćelije koje se aktiviraju lijekovima, sadrže sintetske fosfoantigene i aminobisfosfonate, koji reguliraju endogeni IPP/DMAPP.
Genetičko inženjerstvo
[uredi | uredi izvor]U 2015, tim istraživača predvođen Alexanderom Marsonom[13] na University of California, San Francisco, uspješno je editirao genom ljudskih T-ćelija primjenom Cas9 ribonukleoproteinskog metoda isporuke.[14] Ovaj napredak ima potencijal za primjenu u liječenju putem "immunoterapije raka i terapije na bazi ćelija za HIV, primarnog nedostatka imunosti i autoimunih bolesti.[14]
Poremećaji
[uredi | uredi izvor]Nedostaci
[uredi | uredi izvor]Uzroci nedostataka T-ćelija uključuju limfocitopeniju T-ćelija i/ili nedostatke funkcije pojedinih T-ćelija. Kompletna insuficijencija funkcije T-limfocita može rezultirati zbog nasljednih stanja, kao što su teška kombinirana imunodeficijencija (SCID), Omennov sindrom i hipoplazija hrskavice i kose. Uzroci parcijalnih nedostataka funkcije T-limfocita uključuju sindrom stečene imunodeficijencije (AIDS), i nasljedne promjene, kao što su DiGeorgeov sindrom (DGS), sundrom hromosomskih lomova (HBSD), i B-ćelija i T-ćelija u kombinaciji poremećaja kao što su ataxsija telangiectatica (AT) i Wiskott-Aldrichov sindrom (WAS).
Glavni zabrinjavajući patogeni T-limfocita uključuju unutarćelijske patogene, kao što su i Herpes simplex virus , Mycobacterium i Listeria .[15][16] Pri nedostatku T-ćelija, također su i uobičajene i gljivične infekcije.
Kancer
[uredi | uredi izvor]Kancer T-ćelija se označava limfom T-ćelija, a ubraja se možda kao jedan deset slučajeva ne-Hodkinovog limfoma.[17]
Glavni oblici limfoma T-ćelije su:
- Venčvornio limfom T-ćelija
- Limfom kožnih T-ćelija: Sézaryjev sindrom i Mycosis fungoides
- Anaplastski veliku ćelijski limfom
- Angioimmunoblastbi limfom T-ćelija
Reference
[uredi | uredi izvor]- ^ Alberts B, Johnson A, Lewis J, Raff M, Roberts k, Walter P (2002) Molecular Biology of the Cell. Garland Science: New York, NY pg 1367. "T cells and B cells derive their names from the organs in which they develop. T cells develop in the thymus, and B cells, in mammals, develop in the bone marrow in adults or the liver in fetuses."
- ^ McClory, Susan; Hughes, Tiffany; Freud, Aharon G.; Briercheck, Edward L.; Martin, Chelsea; Trimboli, Anthony J.; Yu, Jianhua; Zhang, Xiaoli; Leone, Gustavo (2012). "Evidence for a stepwise program of extrathymic T cell development within the human tonsil". The Journal of Clinical Investigation. 122 (4): 1403–1415. doi:10.1172/JCI46125. ISSN 1558-8238. PMC 3314444. PMID 22378041.
- ^ Vantourout, Pierre; Hayday, Adrian (2013). "Six-of-the-best: unique contributions of γδ T cells to immunology". Nature Reviews. Immunology. 13 (2): 88–100. doi:10.1038/nri3384. ISSN 1474-1741. PMC 3951794. PMID 23348415.
- ^ Akbar AN, Terry L, Timms A, Beverley PC, Janossy G (april 1988). "Loss of CD45R and gain of UCHL1 reactivity is a feature of primed T cells". J. Immunol. 140 (7): 2171–8. PMID 2965180.CS1 održavanje: upotreba parametra authors (link)
- ^ Willinger T, Freeman T, Hasegawa H, McMichael AJ, Callan MF (2005). "Molecular signatures distinguish human central memory from effector memory CD8 T cell subsets". Journal of Immunology. 175 (9): 5895–903. doi:10.4049/jimmunol.175.9.5895. PMID 16237082.CS1 održavanje: upotreba parametra authors (link)
- ^ Shin, Haina; Iwasaki, Akiko (1. 9. 2013). "Tissue-resident memory T cells". Immunological Reviews. 255 (1): 165–181. doi:10.1111/imr.12087. ISSN 1600-065X. PMC 3748618. PMID 23947354.
- ^ Abbas AK, Benoist C, Bluestone JA, Campbell DJ, Ghosh S, Hori S, Jiang S, Kuchroo VK, Mathis D, Roncarolo MG, Rudensky A, Sakaguchi S, Shevach EM, Vignali DA, Ziegler SF (2013). "Regulatory T cells: recommendations to simplify the nomenclature". Nat. Immunol. 14 (4): 307–8. doi:10.1038/ni.2554. PMID 23507634.CS1 održavanje: upotreba parametra authors (link)
- ^ Singh B, Schwartz JA, Sandrock C, Bellemore SM, Nikoopour E (2013). "Modulation of autoimmune diseases by interleukin (IL)-17 producing regulatory T helper (Th17) cells". Indian J. Med. Res. 138 (5): 591–4. PMC 3928692. PMID 24434314.CS1 održavanje: upotreba parametra authors (link)
- ^ Serriari NE, Eoche M, Lamotte L, Lion J, Fumery M, Marcelo P, Chatelain D, Barre A, Nguyen-Khac E, Lantz O, Dupas JL, Treiner E (2014). "Innate mucosal-associated invariant T (MAIT) cells are activated in inflammatory bowel diseases". Clin. Exp. Immunol. 176: 266–74. doi:10.1111/cei.12277. PMC 3992039. PMID 24450998.CS1 održavanje: upotreba parametra authors (link)
- ^ MR1 antigen presentation to mucosal-associated invariant T cells was highly conserved in evolution. 2009
- ^ Innate T cells detect bacteria. Bacteria, mucosal-associated invariant T cells and MR1. 2010
- ^ Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O'Hair RA, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J (2012). "MR1 presents microbial vitamin B metabolites to MAIT cells". Nature. 491: 717–23. doi:10.1038/nature11605. PMID 23051753.CS1 održavanje: upotreba parametra authors (link)
- ^ http://www.independent.co.uk/news/science/crispr-breakthrough-announced-in-technique-of-editing-dna-to-fight-off-deadly-illnesses-10420050.html
- ^ a b Schumann, Kathrin; Lin, Steven; Boyer, Eric; Simeonov, Dimitre R.; Subramaniam, Meena; Gate, Rachel E.; Haliburton, Genevieve E.; Ye, Chun J.; Bluestone, Jeffrey A. (2015). "Generation of knock-in primary human T cells using Cas9 ribonucleoproteins". Proceedings of the National Academy of Sciences of the United States of America. 112 (33): 10437–10442. doi:10.1073/pnas.1512503112. ISSN 1091-6490. PMC 4547290. PMID 26216948.
- ^ Medscape > T-cell Disorders. Author: Robert A Schwartz, MD, MPH; Chief Editor: Harumi Jyonouchi, MD. Updated: May 16, 2011
- ^ Infection: Microbiology and Management. Wiley-Blackwell. 2006. str. 435. ISBN 1-4051-2665-5. Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) - ^ "The Lymphomas" (PDF). The Leukemia & Lymphoma Society. maj 2006. str. 2. Pristupljeno 7. 4. 2008.
Vanjski linkovi
[uredi | uredi izvor]- Immunobiology, 5th Edition
- niaid.nih.gov – The Immune System
- T-cell Group – Cardiff University
- (Successful!) Treatment of Metastatic Melanoma with Autologous CD4+ T Cells against NY-ESO-1 Arhivirano 5. 1. 2010. na Wayback Machine.
- Davies AJ (1993). "The tale of T cells". Immunology Today. 14 (3): 137–139. doi:10.1016/0167-5699(93)90216-8. PMID 8466629.