Sincicij
| Sincicij | |
|---|---|
Mikrografija dijela poprečnog presjeka skeletnog mišića. Na periferiji svake sincicijske ćelije uočava se nekoliko ćelijskih jedara. | |
| Detalji | |
| Identifikatori | |
| Latinski | Syncytium |
| TH | TH {{{2}}}.html HH1.00.01.0.00020 .{{{2}}}.{{{3}}} |
| Anatomska terminologija | |
Sincicij (grč. σύν - syn = sa, slijedeći + κύτος -kytos = ćelija) – u biologiji – je citoplazmatska masa s više jedara, rezultat fuzije nekoliko ćelija. Termin se nebi trebao miješati sa cenocit (od grčkog κοινός - koinos = zajednički + κύτος - kytos = ćelija), koji je ćeliju koja sadrži mnogo jedara koja nastaje poslije mitoze bez naknadne citokineze. Većina ćelija u eukariotskim organizmima biljaka i životinja imaju jedno jedro; sinciciji su posebni oblici. Međutim, kod nekih organizama to nisu samo uobičajene strukture, već čak predstavljaju najvažnije stanje u životnom ciklusu, kao kod određenih nematoda ili Myxomyceta.
Sincicij se formira fuzijom pojedinih ćelija u slučaju poprečnoprugastih mišića u sisara ili ćeliji nepotpune ćelijske diobe kod insekata, naprimjer. Sincicij je potrebno razlikovati i od plazmodija, što je druga vrsta višejedarne ćelije koja nastaje iz već diferencirane miksamebe ili fuzijom pokretnih grupe gameta Myxomycota.
Funkcija sincicija je raznolika i ovisi o organizmu koji se procjenjuje, mada se općenito može reći da djeluje kao jedinstvena i koordinirana cjelina sastavljena od nekoliko strukturno i funkcionalno povezanih ćelija koje omogućuju razmjenu molekula i električnih impulsa. Sincicij, osim što je rezultat normalnog razvojnog plana organizma, može biti posljedica zaraznih procesa uzrokovanih virusima i drugim patogenima.[1]
Aktivnost mladihi larvi drugog stadija parazitskih nematoda poput rodova Heterodera ili Globodera na konačnom mjestu unutar domaćina uzrokuje zadebljanje ćelijskog zida, zgušnjavanjem citoplazme i spajanjem tri do deset ćelija oko glave parazita. Nastali sincicij ima visoku metaboličku aktivnost, neophodnu za ishranu mladih. Domaćin je, s druge strane, ograničen u funkcionalnosti korijena jer sincicij prekida provodne i drvene sudove. Učinak je smanjenje rasta i prinosa biljaka, napredovanje starenja.[2]
Formiranje
[uredi | uredi izvor]SIncicij se može formirati na dva načina: fuzijom ili dijeljenjem ćelija. Kod jednoćelijskih organizama rjeđi oblik je fuzija citoplazme jedinki iste vrste, bez fuzije jedara. Konjunkcija ovih ameboidnih ćelija naziva se plazmodij. Uobičajen je model kod miksomicetskih gljivica. Fuzija je, s druge strane, oblik zajedničkog sincicija koji omogućava primitivnoj ćeliji da raste uzastopnim djeljenjem jedra, pod cijenu da njihovo segmentiranje ne povećava količinu citoplazme. Ovaj mehanizam može rezultirati multijedarnim jednoćelijskimjedinkama ili postati tvorba višećelijskih organizama. Vrste koje uključuju samo sincicijske organizme uključuju Protozoa, kao i Opalina ranarum i nekoliko vrsta Radiolaria i talofita, među kojima su uključene alge Sifonales i sluzaste gljive.
U višećelijskim organizmima sincicij se može formirati fuzijom ili dijeljenjem ćelija. Mnogi insekti , poput Drosophila melanogaster, polažu jajašca koja se u početku razvijaju kao sincicijski blastoderm, tj. na početku stvaranja embriona imajuju nepotpunu ćelijsku diobu, razdvajaju jedra i ponovno sastavljaju, ali bez citokineze. Zbog toga se jedra množe u zajedničkom citoplazmatskom prostoru. Nasuprot tome, velika vlakna skeletnih mišića formirana su fuzijom pojedinačnih mišićnih ćelija. Mioblasti – ćelije koje potiču iz mišićnih vlakana – prolaze proces diferencijacije usmjeren transkripcijskim faktorima kao što je MyoD, koji ih prisiljava na razmnožavanje u prisustvu faktora rasta. Kad nema IGF-a, mioblasti se prestaju razmnožavati i počinju se spajatii, tvoreći jednu ćeliju s više jedara.
Primjeri
[uredi | uredi izvor]Alge
[uredi | uredi izvor]Nekoliko vrsta algi u potpunosti se sastoji od cenocita. Primjer je rod zelenih algi Caulerpa porodice Caulerpaceae. Jedinke ovih algi čine jedna gigantska ćelija koja može doseći tri metra, što ih čini najvećim živim ćelijama na svijetu.

Ćelije alge reda Chlorophyta su sincicijske zato što ćelijski zid nakon dijeljenja postaje nejasan, kao i ćelija. Suprotno tome, sifonske ili kladoforne alge kao što je Acetabularia započinju život kaojednoćelijsk, ali tada postaju cenocitne.
Talus vrste Siphonocladus pusillus, naprimjer, ima sličan izgled vezikule čiji je ćelijski zid prekriven slojem multijedarne citoplazme, a u centru nalazi se vakuola. Kako jedinka stari, citoplazma se počinje odvajati tvoreći male vezikule s vakuolama prekrivenim slojem polinuklearne citoplazme koji se fragmentira od izvornog sloja. Te male vakuole razvijaju celulozni zid dok lebde unutar središnje vakuole, čiji izgled postaje sličan filamentu sa pregradama koje rastu razgranato bočno, fenomen poznat kao segregacijska dioba i koji je karakterističan za ovu grupu algi. Kod kladoformnih algi, kao što su rodovi Chaetomorpha ili Cladophora, proces je nešto drugačiji. Neovisno o jedarnoj podjeli, citoplazma se podijeli na dva dijela, odvojena od ćelijskog zida, formirajući na tom području membranski prsten prema središtu, a koji stvara dvostruku poprečnu pregradu. Ona se spaja sa svake strane s unutarnjim slojem uzdužnog zida.
Gljive
[uredi | uredi izvor]Većina gljiva rastu kao hife, cilindrične i nitaste strukture promjera 2 do 10 mikrometara i dužine do nekoliko centimetara. Nove hife nastaju obično pojavom novih apeksa duž postojećih hifa procesom zvanim razgranavanje, mada se može dogoditi i da apikalni kraj hifa bifurkira (udvoji), što rezultira u dvije hife s paralelnim rastom. Hife mogu biti pregradne (sa septama) ili cenocitne: prve se dijele u odjeljke s jednim jedrom pomoću unutarnjih pregrada ili septa, koje se formiraju pod pravim uglom prema zidovima hifa. Cenocitne hife se, s druge strane, ne razdvajaju, pa formiraju velike višećelijke jedinstvene strukture.
U mnogih nižih nitastih gljivica micelij je vrlo malo podijeljen i ima malo septi, pa tvori pravi sincicij. Suprotno tome, većina viših vlaknastih gljiva ima dikarion – dva jedra – u kojima su filiformne (nitaste) ćelije dijelom podijeljene u segmente, svaka s dva različita jedra, iz različitih matičnih ćelija, koje se ne spajaju.
Biljke
[uredi | uredi izvor]U procesu dvostruke oplodnje u cvjetnica, jedno od generativnih jedara polenovog zrna dopire do jajne ćelije i nastaje zigot, dok se drugi generativne nukleus veže sa polarnim jedrom centralne ćelije embrionske kese za formiranje triploidnog jedra centralne ćelije ili ćelije iz koje će se razviti endosperm. Mogu biti tri tipa ovog spajanja, ovisno o vrsti procesa. U ćelijskom tipu kariokineza je praćena citokinezama, dakle potiču samo od nepodijeljene ćelije. Suprotno tome, u nuklearnom i helobijalnom tipu, prva podjela endospermogenetskog ćelijskog jedra nije praćena podjelom citoplazme pomoću ćelijskog zida, tako da nastaju cenocitne ćelije koje mogu naknadno formirati ćelijske zidove.[3][4]
U endospermu žitarica, cenocitni stadij razvoja karakterizira sinhronizirana ćelijska podjela oko velike središnje vakuole slijedeći obrazac anteroposteriornog polariteta. Ovaj stadij traje 3 do 5 dana nakon dvostruke oplodnje, ovisno o vrsti i uslovima okoliša, a odmah slijedi niz citokineza koje transformiraju cenocit u višećelijsko tkivo.
Beskičmenjaci
[uredi | uredi izvor]Insekti
[uredi | uredi izvor]
Kod nekoliko vrsta insekata odlučuje se o sudbini matičnih ćelija interakcijom između njihovih različitih dijelova, u procesu koji se naziva sincicijska specifikacija. U embriónu Drosophila melanogaster postoji samo jedna citoplazma za sva jedra. Genetički materijal se razmjenjuje u cijelom jajetu pošto se membrane upravo pojavljuju iz trinaest uzastopnih mitotičkih dioba od njihove oplodnje. Kada sincicij dostigne fazu od oko 1500 ćelija, na vanjskoj strani se pojavljuju pojedinačne monojedarne ćelije, koje će formirati blastoderm.[5][6]
Kičmenjaci
[uredi | uredi izvor]Skeletni mišići
[uredi | uredi izvor]
Za skeletni mišić smatra se ima pravi anatomski sincicij jer svaka ćelija ima više jedara, uzrokovano spajanjem više ćelija. Mioblasti se počinju umnožavati, pod uticajem faktora rasta, poput FGF-a . Receptori integrina iz fibronektina i drugih molekula, kao što su kaderini se poravnavaju a kasnije spajaju u sincicijske mišićne ćelije. Ovaj posljednji proces posreduju meltrini – metaloproteaze. U kulturi, mioblasti formiraju sincicij razmnožavanjem i fuzijom stvarajući kontraktilne proteine. Taj je postupak kooperativan, tako da ćelijska fuzija mijenja medij kultura tako da inducira fuziju drugih mioblasta.
Sincicij skeletnog mišića omogućava brzu koordiniranu kontrakciju mišića po cijeloj njegovoj dužini. Akcijski potencijal propagira se duž površine mišićnih vlakana sa grupom sinaptsnih kontakata s motornog neurona. U patološkim stanjima kao što je miopatija, prisustvo sincicija omogućava vitalnost mišića, jer žarišta nekroze dijela skeletnog mišića ne rezultiraju nekrozom susjednih dijelova tog područja pošto ove regije imaju vlastiti nuklearni materijal, iako neki od preživjelih segmenata ostaju inervirani gubitkom kontinuiteta s neuromuskularni spojem.
Srčani mišić
[uredi | uredi izvor]
Kao i skeletni, srčani mišić je i mišić s prugama zbog prisustva sarkoplazmatskog retikuluma i miofibrila . U početku su istraživači smatrali da su srčana mišićna vlakna sincicij poput skeletnog vlakna. Međutim, s razvojem elektronske mikroskopije utvrđeno je da nije pravi anatomski sincicij jer svako vlakano je odvojena od svojih susjednih sarkolema na bočnom području, koje su kontinuirane sa umetnutim diskovima u kraj svakog od vlakana.
Ovi diskovi imaju jazne raskrsnice sa vrlo visokom provodljivosti, a povezuju citoplazmu susjednih ćelija do dobijanja talasa akcijskog potencijala i pojave depolarizacije – prenesene do kontrakcije pretkomora i komora – bez prisustva neurotransmitera koji pokreću odgovor, tako da se, iako srčani mišić nije anatomski sincicijum, ponaša kao funkcionalni sincicij. Interkalarni diskovi mogu takođe uticati na koheziju srčanog tkiva tokom kontrakcije. U srčanom mišiću proces stvaranja ćelija tkiva sličan je procesu kod Drospphyla melanogaster – podjelom, a ne fuzijom – završavajući ekspresijom miogenina , proteina kojim započinje proces konačne diferencijacije mišićne ćelije.
Glatki mišići
[uredi | uredi izvor]
Za razliku od prugastih skeletnih tkiva, glatkih mišić sadrži slabo razvijena miofibrile razbacane po ćeliji, koji se kontrahiraju zbog razlike u koncentraciji jona kalcija . Glatki mišić može imati jednu ili više jedinica. Iako nijedna od njih nije anatomska sincicij, glatki mišić jednostruke ili višećelijska jedinica može djelovati kao funkcionalni električni sincicij jer su ćelije povezane rascjepom koji omogućuju prolazak živčanog impulsa i koordiniranu kontrakciju, kao odgovor na podsticaj proizveden akcijskim potencijalom. Ovo tkivo je rasprostranjeno u stomaku i probavnim organima, kao što je crijevo. Prisustvo funkcionalnog sincicija u glatkom mišiću koji čini njegove zidove uzrokuje da kad se pobuđuje, stvara se kontraktilni prsten koji pokreće stimulans prema susjednim područjima ispred i iza tačke na kojoj je primljen. Suprotno tome, multijedinični mišići također imaju nehotične kontrakcije, ali neovisno jer su ćelije električno izolirane i stoga nisu sincicij u bilo kojem smislu. Ovo tkivo se može naći na primjer u očnoj šarenici.
Placenta
[uredi | uredi izvor]
Osobito je važan sincicij u kičmenjaka u posteljici sisara. Pretvorba citotrofoblasta – Langhansovog sloja – u sincicij je fiziološki način regeneracije placente. Kada se embrion implantira, okružen je samo citotrofoblastnim elementima. Međutim, neke naučne studije na ljudima pokazale su da 14. dana nakon ovulacije, kada bi trebalo da dođe do menstruacije , ako nema trudnoćae, krvarenje se javlja periovularnom transformacijom u sincicij nezrelog trofoblasta, zbog imunskih reakcija.
Kada se formira posteljica, Langhansov sloj prevladava nad sincicijem. Kako posteljica sazrijeva, odnos se preokreće. Karakteristike distribucije u cirkulaciji krvi fetoplacentnih sudova određuju bolje prokrvljavanje i Langhansovog sloja, kao sincicija, olakšavajući razmjenu između fetusne cirkulacije i međurednom prostoru, koji je prilagođen za bolju oksigenaciju fetusa u trudnoći.
Embrionske ćelije, bez membrana koje ih razdvajaju, tvore višenamjensku barijeru placentnog sincicija koji ima i druge funkcije. U njemu se nalazi veliki broj citoplazmatskih organela, posebno u posljednjim fazama razvoja fetusa, što ukazuje na visoki metabolizam. To je i veliko ležište gvožđa koje se dobija iz majčine plazme. Imunohistohemijskim studijama je takođe pokazano da je sincicij glavno mjesto na kome se formira ljudski horionski gonadotropini.(hCG). Isto tako, kako je otkriveno u tek implantiranom jajetu u kojem trofoblast još nije sazrio, smatra se da potiče i iz citotrofoblasta. Sincicij proizvodi veliku količinu proteina placente koji se raznose u majčinoj krvi. Neki tvrde da izbjegavaju odbacivanje ploda prirodnim imunskim procesima i izbjegavaju zgrušavanje krvi u interventnim prostorima, iako drugi misle da ne ispunjavaju nijednu funkciju. Te proteine oslobađaju otpuštajući faktori koje proizvodi sama placenta, a koja tako djeluje na samostalan način. Tokom trudnoće, proizvodnja proteina konstantno raste.
Kada se trudnoća završi, citotrofoblast se raspada i sincicij postaje izrazito tanak, stvarajući bliski pristup između majčinske i fetusne krvi, do te mjere da ih na nekim mjestima jedva razdvajaju endotel fetusnih kapilara. Ove tačke su poznate kao vaskulo-sincicijske membrane, koje se nalaze ispod sincicija i odvajaju ih od fetusnih kapilara – slično kao i bubrežni glomerul. Ove membrane su specijalizirane za filtriranje krvi; ne nalaze se u nezrelim placentama, niti u zrelim patogenima kod kojih trofoblast nije sazrio. Tako se posteljica može smatrati podeljenom na dvije površine s različitim funkcijama: onu koja odgovara endotelnom –kordijalnom postavljenju – filtrantu, a drugi tvori sincicij sa metaboličkom funkcijom.
Centralni nervni sistem
[uredi | uredi izvor]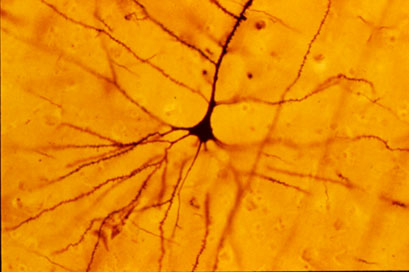

Godine 1855. Franz von Leydig, prvi je predložio da u centralnom živčanom sistema pauka postoji supstanca koju je nazvao Punktsubstanz (=tačkasta supstanca) , a koja se distribuirala u područjima u kojima nije bilo živčanih ćelija. Prema njegovom mišljenju, tvar se sastojala od isprepletenih vlakana. U 1865. Otto Deiters prvi put je vizualizirao produženja nervnih ćelija, iako nije mogao u potpunosti da vidi njihova tijela. Deiters je protumačio da postoje tanki fibrili koji su se protezali od protoplazmatskih ekstenzija i proširili na mijelin, uzrokujući novi aksonski sistema. Sedam godina kasnije, Joseph von Gerlach potvrdio je Leydigovaa i Deiterova zapažanja. Za Gerlacha, nervno tkivo je nastalo kontinuiranim citoplazmatskim sincicijom u kojem su ekstenzije isprepletene i sa anastomozama tvore mrežu. Ta teorija zvala se retikulizam ili retikularna teorija i omogućila je, između ostalog, da se objasni prijenos električnog impulsa između neurona . Njegov i Forel su se suprotstavili ovoj ideji, ali im je nedostajala naučna metodologija koja bi ih podržala.
U 1872. Camillo Golgi razvio je metodu bojenja za analizu mikroskopskih prepatara pripremljenih histološki od nervnog tkiva, koje je danas poznato kao Golgijevo bojenje . Ova tehnika, bazirana na taloženju soli srebra, proizvela je crne siluete ćelija koje, ali njihovi rezultati nisu bili samo u suprotnosti, već skriveni iza strukture nervnih završetaka, što je dovelo do pogrešnog tumačenja. Uprkos tome, uspio je utvrditi da dendriti ne tvore mrežu. Međutim, on je vjerovao da su aksoni izoliranih ćelija to učinili preklapanjem i preplitanjem, omogućujući širenje nervnih impulsa . Kasnije će Santiago Ramón y Cajal poboljšati ovu tehniku razvijajući vlastitu, nazvanu "reducirano srebro" ili Cajal bojenje , koje je omogućila vizualizaciju morfologije neurona. Primjenjujući ove tehnike na embrionskim tkivima i mladim životinjama, Cajal, ne samo da je dokazao da ne postoji sincicij u centralnom nervnom sistemu, već i je razvio modernu teoriju neurona, također objašnjavajući rast nervnih ekstenzija i upuštajući se u proučavanje polarnosti nervnog impulsa.
Uprkos tome, do danas je utvrđeno da neke ćelije koje čine nervni sistema poput astrocita ili nekih džinovskih aksona mogu biti sincicijske. Astrociti izoliraju sinapse drugih susjednih nervnih završetaka i vanćelijsko okruženje. Oni imaju iste jazne veze srčanog mišića, koji se pridružio u klasu sincicije, nazvan mrežni ili glija astrocitni sincicij. Mreža omogućava razmjenu iona i drugih malih molekula, poput neurotransmitera, zbog čega astrociti djeluju kao pufer. Posebna je koncentracija kalijevih iona u nervnom sistemu, pored ostalih metabolita kao što su voda, glutamat i natrijevi ioni, koji čuvaju ionske gradijente, homeostazu i pružaju energetske molekule potrebne za rad neurona.
Ovo je važno u nekim slučajevima, poput hipoksije, kod koje se lokalna koncentracija ovog jona može povećati i do dvadeset puta, depolarizirajući membrane i uzrokujući oslobađanje neurotransmitera koji umnožavaju učinak, dovodeći do smrti neurona. Aksoni mogu biti formirani od jedne ili više ćelija. Unutar višećelijskih aksona nalaze se džinovski aksoni s mijelinom, koji se mogu klasificirati kao segmentirani ili sincicijski. Primjer sincicijskog aksona su džinovski aksoni lignje koji se dobijaju fuzijom od oko tristo do petsto ćelija.[7][8][9][10]
Primjeri patogena
[uredi | uredi izvor]
Nematode
[uredi | uredi izvor]Nekoliko rodova parazitskih nematoda, iz biljaka i životinja, potiče fuziju ćelija domaćina tvoreći sincicij bez kojeg se ne bi mogle prehraniti, razviti ili dovršiti svoj životni ciklus.
Biljni paraziti
[uredi | uredi izvor]Kod nematoda koje formiraju ciste kao Heterodera schachtii, embrionski razvoj je završen unutar sincicija koji se stvara u supstancama vaskularnog parenhima sadržanog u pljuvački larvi drugog stupnja ili J2 koji su prodrli u epidermu korijena sekundarnog domaćina, obično porodica Brassicaceae i Chenopodioideae. Sincicijske ćelije čine hranu parazita i omogućavaju mu da sazre zaštićen cistom. Unutar larve otapa ćelijski zid tvoreći cijevi i praznine bez plazmamembrane, što omogućava jedrima da se kreću i spajaju protoplazme. Od četvrtog stadija J2 razvijaju se odrasle jedinke i izlegu tri do četiri nedelje nakon početne infekcije.
Tod Heterodera i rod parazita krompira, roda Globodera, su sjedeći endoparaziti, ali za razliku od prvog, samo ženke globodere formiraju sincicij. U eksudatnoj tvari kroz korijenje biljaka paraziti podstiču oplodnju i pojavu drugofaznog mladunca, koji prodire u zonu istezanja korijena , u blizini meristema. Larva napreduje stvarajući galerije u korteksu i uzrokujući nekrozu ćelija iz kojih se hrani dok ne dođe do konačnog mjesta ishrane. Larva unosi tvari u korteksne ćelije pored endoderma koji okružuju njenu glavu. Tri do deset ovih ćelija stapaju se jedna s drugom, a njihovi želijski zidovi zadebljavaju, tvoreći sincicij. Kako se larva hrani, ćelijski zidovi se lome i ćelije endoderma postaju nekrotične. Sincicijska citoplazma postaje gušća zbog porasta membranskog materijala, grupiranih ribosoma i organela, iako varira ovisno o kulturi koja se proučava. Povećava metaboličku aktivnost, što je vidljivo u hrapavom endoplazmatskim retikulumima vrlo razvijenim pored mitohondrija, Golgijevog aparata i plastida.[5][6]
Razlika u osjetljivosti kultura na zaraze tim nematodama može biti razlog zašto postoje varijacije u položaju i morfologiji sincitija. Ove činjenice sugeriraju da odnos domaćin-parazit može biti upravljan genima u obje vrste. U kulturama otpornim na nematode, iako se rana faza infekcije razvija na sličan način, krompir stvara unutrašnju reakciju koja tvori tkivo nalik sincicija, ali okruženo membranom koja je vjerovatno sastavljena od nekrotičnih ćelija koja ne dopušta J2 da sazrije, uslijed liziranja ćelijskog zida i citoplazme sincicija, što smanjuje energiju dostupnu larvama. Takođe mogu imati i amiloplaste i mitohondrije koje mogu neutralizirati proizvode koje izlučuju paraziti i smanjiti količinu hrane na raspolaganju za njihovu ishranu. Postoje studije koje ukazuju na to da aminokiseline lizin, fenilalanin, triptofan i posebno metionin imaju štetne učinke kako na metabolizam sincijecija, tako i na razvoj same nematode.
Životinjski paraziti
[uredi | uredi izvor]Različite podvrste roda Trichinella izazivaju zoonozu zvanu trihineloza, kojom parazit kolonizira prugasto mišićno tkivo kod sisara . Nositelj infekcije je unos mesa životinja koje su zaražene cistama nematoda. Izuvijana larva formira spiralu unutar ciste. Gutanjem ciste enzimi za probavu oslobađaju larvu. Ovo se prenosi pasivno u tankom crijrvu,a putem peristaltike dok ne dostigne fazu odraslih. Tamo se razmnožavaju, a novo leglo larvi prožima crijevnu sluznicu i zaštićeni su formiranjem sincicija fuzijom približnog broja od sto dvadeset ćelija. U unutrašnjosti prolaze kroz četiri promjene dok završe svoj razvoj između dan i dan i po kasnije. Larve se prenose putem linfno – krvnih sudova i parazitirau izbrazdanim skeletnim mišićima.
Ljudi su povremeni domaćini, često prenošenjem putem unosa zaražene svinjetine. Vrsta koja nanosi najviše štete ljudima je Trichinella spiralis.
Virusi
[uredi | uredi izvor]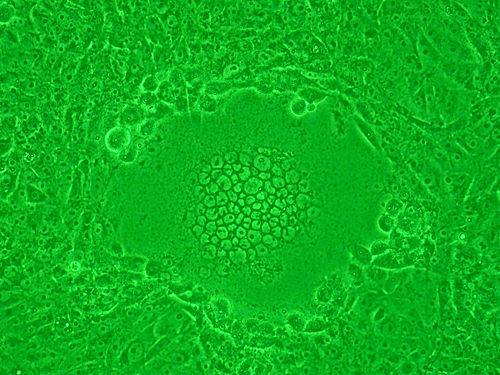
Određene vrste virusa, poput onih u porodici Paramyxoviridae – uključujući respiratorni sincicijski virus (RSV) i HIV – također formiraju sincicij, ali sa ćelijama domaćina . Goveđi respiratorni sincicijski virus (BRSV) nije prekriven cilija ćelija bronhijskog epitela i alveola razmnožavaju se formiranjem velike sincicijske ćelije sa akumulacijom limfocita u intersticijalnom tkivu. U sarem polovine novorođenčadi hospitalizirane zbog bronhiolitisa u Sjedinjenim Državama etiološki agens je bio respiratorni sincicijski virus. Virus izaziva sincicij u disajnim putevima bebe, koji vjerovatno izaziva reakciju na imunoglobulin E i tvari koje stvaraju neutrofili, što uzrokuje edem u sluznici, bronhospazam i na kraju nekroze, od epitela od respiratornog sistema, jer ogromne ćelije sincicija začepljuju alveole, bronhiole i bronhije, otežavajući disanje.[11][12]
Kod HIV infekcije, virus inficira pomoćnu T-ćeliju. Tada ćelija počinje da ispoljava površinske glikoproteine HIV-a koji su antigeni . Tipski, citotoksični T-limfocit odmah počinje ubrizgavati limfotoksin, kao perforin ili granzime koji ubijaju zaražene pomoćne T-ćelije. Međutim, ako u blizini postoje sursdničkr T-ćrlije, receptori za HIV gp41 koji se pojavljuju na površini T-ćelije vežu se na druge slične limfocite. To uzrokuje stapanje na desetine suradničkih preko ćelijskih membrana u T-ćelijama džinovskog nefunkcionalnofg sincicija, zbog kojeg virion HIV-a može ubiti mnoge pomoćne T-ćelije, inficirajući samo jednu.
Infekcija HIV-1 utiče na normalan metabolizam lipida kod zaraženih pacijenata. Kako se bolest pogoršava. povećava koncentraciju triglicerid]a i smanjuje polunezasićene masne kiseline i holesterol u krvi. Konkretno, nizak nivo holesterola i HDL mogao bi doprinijeti stvaranju sincicija i izazvati ekspresiju fenotipa HIV-a. Provedene su in vitro studije koje sugeriraju da HIV stvara veliku desaturaciju [[[stearinska kiselina|stearinske kiseline]], čineći je oleinskom, mijenjajući prirodni odnos između njih u ćelijskim membranama, što pomažw stvaranju sincicija. Čini se da je to također povezano i sa iscrpljivanjem ćelija CD4. Toom ćelijske fuzije dolazi do metaboličkih promjena u ćelijama domaćina, koje desaturiraju masne kiseline lipidnog dvosloja, mijenjajući njihovu fluidnost. To pridonosi stvaranju sincicija i pogodovalo ekspresiji virusnih fenotipa. Ova ideja je pojačana jer njeno formiranje inhibira apolipoprotein Al, glavni u HDL.[13][14]
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Daubenmire, R. F. (1936). "The Use of the Terms Coenocyte and Syncytium in Biology". Science. 84 (2189): 533–534. doi:10.1126/science.84.2189.533. PMID 17806555.
- ^ Willmer, P. G. (1990). Invertebrate Relationships: Patterns in Animal Evolution. Cambridge University Press, Cambridge.
- ^ Tiwari, S. C.; Gunning, B. E. S. (1986). "Colchicine inhibits plasmodium formation and disrupts pathways of sporopollenin secretion in the anther tapetum ofTradescantia virginiana L". Protoplasma. 133 (2–3): 115. doi:10.1007/BF01304627.
- ^ Murguía-Sánchez, G. (2002). "Embryo sac development in Vanroyenella plumosa, Podostemaceae". Aquatic Botany. 73 (3): 201–210. doi:10.1016/S0304-3770(02)00025-6.
- ^ a b Gobert, Geoffrey N.; Stenzel, Deborah J.; McManus, Donald P.; Jones, Malcolm K. (decembar 2003). "The ultrastructural architecture of the adult Schistosoma japonicum tegument". International Journal for Parasitology. 33 (14): 1561–1575. doi:10.1016/s0020-7519(03)00255-8. ISSN 0020-7519. PMID 14636672.
- ^ a b Jerome), Bogitsh, Burton J. (Burton (2005). Human parasitology. Carter, Clint E. (Clint Earl), Oeltmann, Thomas N. Burlington, MA: Elsevier Academic Press. ISBN 0120884682. OCLC 769187741.
- ^ http://www.naturalezadearagon.com/historianatural/biologiageneral3.php%7Ctitle= Biología General. La Célula: Citología. Los seres pluricelulares.
- ^ Histología y Embriología del Ser Humano/ Histology and Embryology of the Human Being: Bases Celulares Y Moleculares/ Cellular and Molecular Basis. Nepoznati parametar
|editors=zanemaren (prijedlog zamjene:|editor=) (pomoć) - ^ http://www.medicoscubanos.com/
- ^ http://books.google.com.uy/books?id=NxYmIRZQi2oC&pg=PA321&dq=diferenciaci%C3%B3n+de+mioblasto&hl=es&ei=oU3dTYz-Gonu0gGK-fXwDw&sa=X&oi=book_result&ct=result&resnum=5&ved=0CDwQ6AEwBDgK#v=onepage&q&f=false
- ^ Albrecht, Thomas; Fons, Michael; Boldogh, Istvan; Rabson, Alan S. (1. 1. 1996). Baron, Samuel (ured.). Medical Microbiology (4th izd.). Galveston (TX): University of Texas Medical Branch at Galveston. ISBN 0963117211. PMID 21413282.
- ^ "ViralZone: Syncytium formation induced by viral infection". viralzone.expasy.org. Arhivirano s originala, 21. 12. 2016. Pristupljeno 16. 12. 2016.
- ^ Salsman, Jayme; Top, Deniz; Boutilier, Julie; Duncan, Roy (1. 7. 2005). "Extensive Syncytium Formation Mediated by the Reovirus FAST Proteins Triggers Apoptosis-Induced Membrane Instability". Journal of Virology (jezik: engleski). 79 (13): 8090–8100. doi:10.1128/JVI.79.13.8090-8100.2005. ISSN 0022-538X. PMC 1143762. PMID 15956554.
- ^ Duncan, Roy; Corcoran, Jennifer; Shou, Jingyun; Stoltz, Don (5. 2. 2004). "Reptilian reovirus: a new fusogenic orthoreovirus species". Virology. 319 (1): 131–140. doi:10.1016/j.virol.2003.10.025. PMID 14967494.