Бялкі

Бялкі (пратэіны, поліпептыды) — высокамалекулярныя арганічныя рэчывы, якія складаюцца са звязаных паміж сабой у ланцуг пептыднай сувяззю амінакіслотаў. Адзін з асноўных хімічных кампанентаў абмену рэчываў і энергіі жывых арганізмаў. Абумоўліваюць іх будову, галоўныя адзнакі, функцыі, разнастайнасць і адаптацыйныя магчымасці, удзельнічаюць ва ўтварэнні клетак, тканак і органаў (структурныя бялкі), у рэгуляцыі абмену рэчываў (гармоны), з'яўляюцца запасным пажыўным рэчывам (запасныя бялкі).
З двух дзясяткаў амінакіслот пабудаваны малекулы многіх тысяч розных бялкоў. Гэта робіцца магчымым дзякуючы рознай паслядоўнасці злучэння амінакіслотных астаткаў у макрамалекуле бялку. Кожны бялок мае строга пэўную паслядоўнасць і лік амінакіслотнах астаткаў.
Бялкі складаюць матэрыяльную аснову амаль усіх жыццёвых працэсаў: росту, стрававання, размнажэння, ахоўных функцый арганізма, утварэння генетычнага апарату і перадачы спадчынных прыкмет (нуклеапратэіды), пераносу ў арганізме рэчываў (транспартныя бялкі), скарачэння мышцаў, перадачы нервовых імпульсаў і інш. Ферменты бялковай прыроды выконваюць у арганізме спецыфічныя каталітычныя функцыі, выключна важнае значэнне ў рэгуляцыі фізіялагічных працэсаў маюць бялкі-гармоны.
Сінтэзуюцца бялкі з неарганічных рэчываў раслінамі і некаторымі бактэрыямі. Жывёлы і чалавек атрымліваюць гатовыя бялкі з ежы. З прадуктаў іх расшчаплення (пептыдаў і амінакіслот) у арганізме сінтэзуюцца спецыфічныя ўласныя бялкі, дзе яны няспынна разбураюцца і замяняюцца зноў сінтэзаванымі. Біясінтэз бялкоў ажыццяўляецца па матрычным прынцыпе з удзелам ДНК, РНК, пераважна ў рыбасомах клетак і інш. Паслядоўнасць амінакіслот у бялках адлюстроўвае паслядоўнасць нуклеатыдаў у нуклеінавых кіслотах.
Гісторыя вывучэння
[правіць | правіць зыходнік]Бялкі былі выдзелены ў асобны клас біялагічных малекул у 18 стагоддзі ў выніку работ французскага хіміка Антуана дэ Фуркруа і іншых вучоных, у якіх была адзначана ўласцівасць бялкоў каагуляваць (дэнатураваць) пад уздзеяннем награвання або кіслот. У той час былі даследаваны такія бялкі, як альбумін («яечны бялок»), фібрын (бялок з крыві) і глютэн са збожжа пшаніцы.

У пачатку 19 стагоддзя ўжо былі атрыманы некаторыя звесткі аб элементарным складзе бялкоў, было вядома, што пры гідролізе бялкоў ўтвараюцца амінакіслоты. Некаторыя з гэтых амінакіслот (напрыклад, гліцын і лейцын) ужо былі ахарактарызаваны. Галандскі хімік Герыт Мульдэр на аснове аналізу хімічнага складу бялкоў прапанаваў гіпотэзу, што амаль усе бялкі маюць падобную эмпірычную формулу. У 1836 годзе Мульдэр прапанаваў першую мадэль хімічнай будовы бялкоў. Грунтуючыся на тэорыі радыкалаў, ён пасля некалькіх удакладненняў прыйшоў да высновы, што мінімальная структурная адзінка бялку валодае наступным складам: C40H62N10O12. Гэтую адзінку ён назваў «пратэінам» (Pr) (ад грэч. протас — першы, першасны), а тэорыю — «тэорыяй пратэіна»[1].
Сам тэрмін «пратэін» быў прапанаваны яшчэ шведскім хімікам Якабам Берцэліусам[2]. Згодна з уяўленнямі Мульдэра, кожны бялок складаецца з некалькіх пратэінавых адзінак, серы і фосфару. Напрыклад, ён прапанаваў запісваць формулу фібрына як 10PrSP. Мульдэр таксама даследаваў прадукты разбурэння бялкоў — амінакіслоты і для адной з іх (лейцын) з малой доляй памылкі вызначыў малекулярную масу — 131 дальтон. Па меры назапашвання новых дадзеных пра бялкі тэорыя пратэіна стала падвяргацца крытыцы, але, нягледзячы на гэта, да канца 1850-х усё яшчэ лічылася агульнапрызнанай.
Да канца 19 стагоддзя было даследавана большасць амінакіслот, якія ўваходзяць у склад бялкоў. У канцы 1880-х гадоў рускі вучоны А. Я. Данілеўскі адзначыў існаванне пептыдных груп (CO-NH) у малекуле бялку [3][4]. У 1894 годзе нямецкі фізіёлаг Альбрэхт Косель вылучыў тэорыю, згодна з якой менавіта амінакіслоты з'яўляюцца асноўнымі структурнымі элементамі бялкоў[5]. У пачатку 20 стагоддзя нямецкі хімік Эміль Фішэр эксперыментальна даказаў, што бялкі складаюцца з амінакіслотных астаткаў, злучаных пептыднымі сувязямі. Ён жа ажыццявіў першы аналіз амінакіслотнай паслядоўнасці бялку і патлумачыў з'яву пратэоліза.
Аднак цэнтральная роля бялкоў у арганізме не была прызнана да 1926 года, калі амерыканскі хімік Джэймс Самнер (пасля — лаўрэат Нобелеўскай прэміі па хіміі) паказаў, што фермент урэазы з'яўляецца бялком [6].
Складанасць выдзялення чыстых бялкоў абцяжарвала іх вывучэнне. Таму першыя даследаванні праводзіліся з выкарыстаннем тых поліпептыдаў, якія лёгка маглі быць ачышчаны ў вялікай колькасці, гэта значыць бялкоў крыві, курыных яек, розных таксінаў, а таксама стрававальных / метабалічных ферментаў, што выдзяляюцца пасля забою жывёлы. У канцы 1950-х гадоў кампанія Armour Hot Dog Co. змагла ачысціць кілаграм бычынай панкрэатычнай рыбануклеазы А, якая стала эксперыментальным аб'ектам для шматлікіх даследаванняў.
Ідэя аб тым, што другасная структура бялкоў — вынік утварэння вадародных сувязяў паміж амінакіслотнымі астаткамі, была выказана Уільямам Астберы ў 1933 годзе, але Лайнус Карл Полінг лічыцца першым вучоным, які змог паспяхова прадказаць другасную структуру бялкоў. Пазней Уолтар Каўзман, абапіраючыся на працы Кая Ліндэрстром-Ланга, унёс значны ўклад у разуменне законаў утварэння трацічнай структуры бялкоў і ролі ў гэтым працэсе гідрафобных узаемадзеянняў. У канцы 1940-х — пачатку 1950-х гадоў Фрэдэрык Сенгер распрацаваў метад секвеніравання бялкоў, з дапамогай якога ён да 1955 года вызначыў амінакіслотную паслядоўнасць двух ланцугоў інсуліну[7][8][9], прадэманстраваўшы, што бялкі — гэта лінейныя палімеры амінакіслот, а не разгалінаваныя (як у некаторых цукроў) ланцугі, калаіды або цыклолы.
Першыя прасторавыя структуры бялкоў, атрыманыя метадам дыфракцыі рэнтгенаўскіх прамянёў (рэнтгенаструктурнага аналізу) сталі вядомы ў канцы 1950-х — пачатку 1960-х гадоў, а структуры, адкрытыя з дапамогай ядзернага магнітнага рэзанансу — у 1980-х гадах. У 2012 годзе Банк дадзеных аб бялках (Protein Data Bank) утрымліваў каля 87 000 структур бялкоў [10].
У 21 стагоддзі даследаванне бялкоў перайшло на якасна новы ўзровень, калі даследуюцца не толькі індывідуальныя ачышчаныя бялкі, але і адначасовае змяненне колькасці і посттрансляцыённых мадыфікацый вялікай колькасці бялкоў асобных клетак, тканак або цэлых арганізмаў. Гэтая галіна біяхіміі называецца пратэёміка. З дапамогай метадаў біяінфарматыкі стала магчыма не толькі апрацоўваць дадзеныя рэнтгенаструктурнага аналізу, але і прадказваць структуру бялку на падставе яго амінакіслотнай паслядоўнасці. У цяперашні час крыаэлектронная мікраскапія буйных бялковых комплексаў і прадказанне прасторавых структур бялковых даменаў з дапамогай камп'ютарных праграм набліжаюцца да атамарнай дакладнасці[11].
Класіфікацыя
[правіць | правіць зыходнік]Паводле паходжання і крыніц атрымання бялкі падзяляюцца на
- раслінныя,
- жывёльныя,
- бактэрыяльныя.
Паводле хімічнага саставу выдзяляюць бялкі
- простыя (некан'югіраваныя) — пратэіны, якія складаюцца з астаткаў амінакіслот, што злучаны паміж сабою пептыднай сувяззю (—NH—CO) у доўгія ланцугі — поліпептыды;
- складаныя (кан'югіраваныя) — пратэіды, якія складюцца з простага бялка, злучанага з небялковым арганічным ці неарганічным кампанентам непептыднай прыроды, т. зв. прастэтычнай групай, далучанай да поліпептыднай часткі. Сярод складаных бялкоў паводле тыпу прастэтычнай групы вылучаюць
- нуклеапратэіды
- фосфапратэіды
- глікапратэіды
- металапратэіды
- гемапратэіды
- флавапратэіды
- ліпапратэіды і інш.
Уласцівасці
[правіць | правіць зыходнік]
Памер
[правіць | правіць зыходнік]Памер бялку можа вымярацца колькасцю амінакіслотных астаткаў або ў дальтонах (малекулярная маса), але з-за адносна вялікай велічыні малекулы маса бялку вымяраецца ў вытворных адзінках — кіладальтонах (кДа). Бялкі дрожджаў, у сярэднім, складаюцца з 466 амінакіслотных астаткаў і маюць малекулярную масу 53 кДа. Самы вялікі з вядомых у цяперашні час бялкоў — тытын — з'яўляецца кампанентам саркамераў мускулаў; малекулярная маса яго розных варыянтаў (ізаформ) вар'іруецца ад 3000 да 3700 кДа. Тытын камбалападобнай цягліцы (лац.: soleus) чалавека складаецца з 38138 амінакіслот[12].
Для вызначэння малекулярнай масы бялкоў выкарыстоўваюць такія метады, як гель-фільтраванне, электрафарэз у поліакріламіднам гелі, мас-спектраметрычны аналіз, седыментацыйны аналіз і іншыя[13].
Фізіка-хімічныя ўласцівасці
[правіць | правіць зыходнік]Агульны хімічны састаў бялкоў (у % у пераліку на сухое рэчыва): вуглярод — 50-55, кісларод — 21-23, азот — 15-18, вадарод — 6-7,5, сера — 0,3-2,5, фосфар — 1-2 і інш.
Большасць бялкоў раствараецца ў вадзе і ўтварае малекулярныя растворы.
Па растваральнасці ў вадзе, растворах нейтральных соляў, шчолачаў, кіслотах і арганічных растваральніках вылучаюць альбуміны, гістоны, глабуліны, глютэліны, праламіны, пратаміны і пратэіноіды.
Бялкі маюць кіслыя карбаксільныя і амінныя групы, таму ў растворах яны амфатэрныя (маюць уласцівасці асноў і кіслот). Пры гідролізе яны распадаюцца да амінакіслот; пад уплывам розных фактараў здольныя да дэнатурацыі і каагуляцыі, уступаюць у рэакцыі акіслення, аднаўлення, нітравання і інш.
Пры пэўных значэннях pH у растворах бялкоў пераважае дысацыяцыя тых ці іншых груп, што надае ім адпаведны зарад і выклікае рух у электрычным полі — электрафарэз.
Структура
[правіць | правіць зыходнік]Структура бялкоў характарызуецца амінакіслотным саставам, парадкам чаргавання амінакіслотных астаткаў у поліпептыдных ланцугах, іх даўжынёй і размеркаваннем у прасторы.
Амінакілотны састаў
[правіць | правіць зыходнік]У састаў бялкоў уваходзіць ад 50 да 6000 і больш астаткаў 20 амінакіслот, што ўтвараюць складаныя поліпептыдныя ланцугі.
Амінакіслотны састаў розных бялкоў неаднолькавы і з'яўляецца іх істотнай характарыстыкай, а таксама мерай харчовай каштоўнасці. Паслядоўнасць амінакіслот у кожным бялку вызначаецца паслядоўнасцю монануклеатыдных будаўнічых блокаў у асобных адрэзках малекулы ДНК. Вядома амінакіслотная паслядоўнасць некалькіх соцень бялкоў (напрыклад, адрэнакортыкатропнага гармону чалавека, рыбануклеазы, цытахромаў, гемаглабіну і інш.).
Парушэнні амінакіслотнай паслядоўнасці ў малекуле бялка выклікаюць т.зв. малекулярныя хваробы.
Амінакіслотную паслядоўнасць поліпептыднага ланцуга для малекулы гармону інсуліну ўстанавіў у 1953 годзе англійскі біяхімік Ф. Сенгер.
Звесткі пра колькасць адрозненняў у амінакіслотных паслядоўнасцях гамалагічных бялкоў, узятых з розных відаў арганізмаў, выкарыстоўваюць пры складанні эвалюцыйных картаў, якія адлюстроўваюць паслядоўныя этапы ўзнікнення і развіцця пэўных відаў арганізмаў у працэсе эвалюцыі.
Узроўні арганізацыі
[правіць | правіць зыходнік]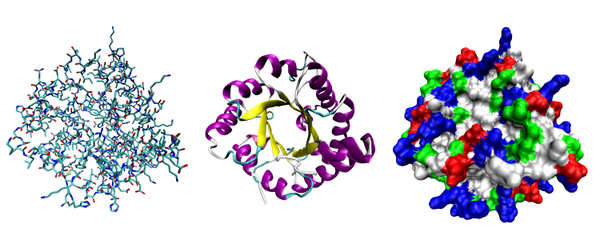
К. Ліндстром-Ланг прапанаваў выдзяляць 4 ўзроўні структурнай арганізацыі бялкоў: першасную, другасную, трацічную і чацвярцічную структуры. Хоць такі падзел крыху састарэў, ім працягваюць карыстацца[1]. Першасная структура (паслядоўнасць амінакіслотных астаткаў) поліпептыда вызначаецца структурай яго гена і генетычным кодам, а структуры больш высокіх парадкаў фарміруюцца ў працэсе згортвання бялку[14]. Хоць прасторавая структура бялку ў цэлым вызначаецца яго амінакіслотнай паслядоўнасцю, яна з'яўляецца даволі лабільнай і можа залежыць ад знешніх умоў, таму больш правільна казаць пра пераважную або найбольш энергетычна выгадную канфармацыю бялку[1].
4 парадкі (узроўні) структуры бялкоў:
- першасная (лінейная паслядоўнасць амінакіслотных астаткаў у поліпептыдным ланцугу),
- другасная (прасторавая, найчасцей спіральная прасторавая канфігурацыя, якую прымае сам поліпептыдны ланцуг),
- трацічная (трохмерная канфігурацыя, якая ўзнікае ў выніку складвання або закручвання структур другаснага парадку ў больш кампактную глабулярную форму),
- чацвярцічная (злучэнне некалькіх частак з трацічнай структурай у адну больш буйную комплексную праз некавалентныя сувязі).
Найбольш устойлівая першасная структура бялкоў, іншыя лёгка разбураюцца пры павышэнні тэмпературы, рэзкім змяненні pH асяроддзя і іншых уздзеяннях (дэнатурацыя бялкоў), што вядзе да страты асноўных біялагічных уласцівасцяў. Фарміраванне прасторавай канфігурацыі малекул бялка вызначаецца наяўнасцю ў поліпептыдных ланцугах вадародных, дысульфідных, эфірных і салявых сувязяў, сіл Ван дэр Ваальса і інш.
Класіфікацыя па тыпу будовы
[правіць | правіць зыходнік]Па агульнаму тыпу будовы бялкі можна разбіць на тры групы:
- Фібрылярныя бялкі — утвараюць палімеры, іх структура звычайна высокарэгулярна і падтрымліваецца, у асноўным, узаемадзеяннямі паміж рознымі ланцугамі. Яны ўтвараюць мікрафіламенты, мікратрубачкі, фібрылы, падтрымліваюць структуру клетак і тканак. Да фібрылярных бялкоў адносяцца кератын і калаген.
- Глабулярныя бялкі — вадарастваральныя, агульная форма малекулы больш ці менш сферычная.
- Мембранныя бялкі — маюць перасякаючыя клеткавую мембрану дамены, але часткі іх выступаюць з мембраны ў міжклеткавае асяроддзе і цытаплазму клеткі. Мембранныя бялкі выконваюць функцыю рэцэптараў, гэта значыць ажыццяўляюць перадачу сігналаў, а таксама забяспечваюць трансмембранны транспарт розных рэчываў. Бялкі-транспарцёры спецыфічныя, кожны з іх прапускае праз мембрану толькі пэўныя малекулы або пэўны тып сігналу.
Гаспадарчае значэнне
[правіць | правіць зыходнік]Многія прыродныя бялкі і бялковыя ўтварэнні выкарыстоўваюць у прамысловасці (напрыклад, для вырабу скуры, шэрсці, натуральнага шоўку, казеіну, пластмасаў і інш.), медыцыне і ветэрынарыі (як лекавыя сродкі і біястымулятары, напрыклад, інсулін пры цукровым дыябеце, сываратачны альбумін як заменнік крыві, гама-глабулін для прафілактыкі інфекцыйных захворванняў, бялкі-ферменты для лячэння парушэнняў абмену рэчываў, гідралізатары бялкоў для штучнага жыўлення).
Для атрымання пажыўных і кармавых бялкоў выкарыстоўваюць мікрабіялагічны сінтэз. Вядуцца даследаванні па штучным сінтэзе бялковых малекул (штучна сінтэзаваны фермент рыбануклеаза і інш.).
Бялкі — адзін з галоўных аб'ектаў даследаванняў біяхіміі, імуналогіі і іншых раздзелаў біялагічнай навукі.
Посттрасляцыйныя мадыфікацыя
[правіць | правіць зыходнік]Пасля завяршэння трансляцыі большасць бялкоў падвяргаецца далейшым хімічным мадыфікацыям, якія называюцца посттрасляцыйнымі мадыфікацыямі[15]. Вядома больш за дзвесце варыянтаў посттрасляцыйных мадыфікацый бялкоў[16].
Посттрасляцыйныя мадыфікацыі могуць рэгуліраваць працягласць існавання бялкоў у клетцы, іх ферментатыўную актыўнасць і ўзаемадзеянне з іншымі бялкамі. У шэрагу выпадкаў посттрасляцыйныя мадыфікацыі з'яўляюцца абавязковым этапам паспявання бялку, у адваротным выпадку ён аказваецца функцыянальна неактыўным. Напрыклад, пры паспяванні інсуліну і некаторых іншых гармонаў неабходны абмежаваны пратэоліз поліпептыднага ланцуга, а пры паспяванні бялкоў плазматычнай мембраны — гліказіліраванне.
Посттрасляцыйныя мадыфікацыі могуць быць як шырока распаўсюджанымі, так і рэдкімі, нават унікальнымі. Прыкладам універсальнай мадыфікацыі служыць убіквіцініраванне (далучэнне да бялку ланцуга з некалькіх малекул кароткага бялку убіквіціна), якое служыць сігналам да расшчаплення гэтага бялку пратэасомай[17]. Іншай распаўсюджанай мадыфікацыяй з'яўляецца гліказіліраванне — лічыцца, што каля паловы бялкоў чалавека гліказіліраваныя[18]. Да рэдкіх мадыфікацый адносяць тыразініраванне/дэтыразініраванне і полігліцыліраванне тубуліна[19].
Адзін і той жа бялок можа падвяргацца шматлікім мадыфікацыям. Так, гістоны (бялкі, якія ўваходзяць у склад храмаціна ў эўкарыётах) у розных умовах могуць падвяргацца больш чым 150 розным мадыфікацыям[20].
Посттрасляцыйныя мадыфікацыі дзеляць на:
- мадыфікацыі галоўнага ланцугу;
- адшчапленнем N-канцавога астатку метыаніна;
- абмежаваны пратэоліз — выдаленне фрагмента бялку, якое можа адбывацца з канцоў (адшчапленнем сігнальных паслядоўнасцей) або, у асобных выпадках, у сярэдзіне малекулы (паспяванне інсуліну);
- далучэнне розных хімічных груп да свабодных аміна- і карбаксільных груп (N-ацыліраванне, мірыстаіліраванне і інш.);
- мадыфікацыі бакавых ланцугоў амінакіслот;
- далучэнне або адшчапленне невялікіх хімічных груп (гліказіліраванне, фасфараліраванне і інш.);
- далучэнне ліпідаў і вуглевадародаў;
- змена стандартных амінакіслотных астаткаў на нестандартныя (утварэнне цытруліна);
- утварэнне дысульфідных масткоў паміж рэшткамі цыстэіна;
- далучэнне невялікіх бялкоў (сумаіліраванне і убіквіцініраванне).
Зноскі
- ↑ а б в Ю. А. Овчинников. Биоорганическая химия. — Москва: Просвещение, 1987. — С. 24—26.
- ↑ Henry Leicester. Berzelius, Jöns Jacob // Dictionary of Scientific Biography 2. — New York: Charles Scribner’s Sons, 1980. — С. 90—97. — ISBN 0-684-10114-9.
- ↑ Данилевский А.Я. Биолого-химические сообщения о белковых веществах (материалы для химической конституции и биогенеза их) // Физиологический сборник. — 1888. — Т. 1. — С. 289.
- ↑ Цветков Л. А. § 38. Белки // Органическая химия. Учебник для 10 класса. — 20-е изд. — М.: Просвещение, 1981. — С. 184—193. — 1 210 000 экз.
- ↑ Белки // Химическая энциклопедия. — Москва: Советская энциклопедия, 1988.
- ↑ N. H. Barton, D. E. G. Briggs, J. A. Eisen. Evolution. — Cold Spring Harbor Laboratory Press, 2007. — С. 38. — ISBN 978-0-87969-684-9.
- ↑ Нобелеўская лекцыя Ф. Сенгера. Архівавана з першакрыніцы 5 студзеня 2013. Праверана 3 студзеня 2013.
- ↑ Sanger F., Tuppy H. The amino-acid sequence in the phenylalanyl chain of insulin. 2. The investigation of peptides from enzymic hydrolysates // Biochem J. — 1951. — В. 4. — Т. 49. — С. 481—490. — PMID 14886311.
- ↑ Sanger F., Thompson E. O. The amino-acid sequence in the glycyl chain of insulin. II. The investigation of peptides from enzymic hydrolysates // Biochem J. — 1953. — В. 3. — Т. 53. — С. 366—374. — PMID 13032079.
- ↑ Protein Data Bank(недаступная спасылка). Rutgers and UCSD. — Biological Macromolecular Resource. Архівавана з першакрыніцы 27 снежня 2012. Праверана 26 снежня 2012.
- ↑ Yahav T., Maimon T., Grossman E., Dahan I., Medalia O. Cryo-electron tomography: gaining insight into cellular processes by structural approaches // Curr Opin Struct Biol. — 2011. — В. 5. — Т. 21. — С. 670—677. — PMID 21813274.
- ↑ Fulton A., Isaacs W. Titin, a huge, elastic sarcomeric protein with a probable role in morphogenesis // Bioessays. — 1991. — В. 4. — Т. 13. — С. 157—161. — PMID 1859393.
- ↑ Х.-Д. Якубке, Х. Ешкайт. Глава 3.5 Физико-химические свойства // Аминокислоты, пептиды, белки. — Москва: Мир, 1985. — С. 356—363.
- ↑ Ленинджер А. Основы биохимии в 3 томах. — Москва: Мир, 1985.
- ↑ Jones D. T. Protein secondary structure prediction based on position-specific scoring matrices // J Mol Biol. — 1999. — В. 2. — Т. 292. — С. 195—202. — PMID 10493868.
- ↑ Jensen O. N. Interpreting the protein language using proteomics // Nat Rev Mol Cell Biol. — 2006. — В. 6. — Т. 7. — С. 391—403. — PMID 16723975.
- ↑ Demartino G. N., Gillette T. G. Proteasomes: machines for all reasons // Cell. — 2007. — В. 4. — Т. 129. — С. 659—662. — PMID 17512401.
- ↑ Walsh G., Jefferis R. Post-translational modifications in the context of therapeutic proteins // Nat Biotechnol. — 2006. — В. 10. — Т. 24. — С. 1241—1252. — PMID 17033665.
- ↑ Rosenbaum, J. (2000). "Cytoskeleton: functions for tubulin modifications at last". Curr Biol. 10: 801–803. doi:10.1016/S0960-9822(00)00767-3. ISSN 0960-9822. PMID 11084355.
- ↑ Bronner C., Chataigneau T., Schini-Kerth V. B., Landry Y. The "Epigenetic Code Replication Machinery", ECREM: a promising drugable target of the epigenetic cell memory // Curr Med Chem. — 2007. — В. 25. — Т. 14. — С. 2629—2641. — PMID 17979715.
Літаратура
[правіць | правіць зыходнік]- Беларуская энцыклапедыя: У 18 т. Т. 3: Беларусы — Варанец / Рэдкал.: Г. П. Пашкоў і інш. — Мн. : БелЭн, 1996. — Т. 3. — 511 с. — 10 000 экз. — ISBN 985-11-0035-8. — ISBN 985-11-0068-4 (т. 3).
Спасылкі
[правіць | правіць зыходнік]На Вікісховішчы ёсць медыяфайлы па тэме Бялкі
Слоўнікі і энцыклапедыі | Вялікая каталанская · Вялікая кітайская · Вялікая кітайская · Вялікая нарвежская · Вялікая расійская (старая версія) · Вялікая савецкая (1 выд.) · Бракгаўза і Эфрона · Кругасвет · Ларуса · Малы Бракгаўза і Эфрона · Britannica (онлайн) · Brockhaus · Brockhaus · Treccani · Universalis · Сучаснай Украіны | |||
|---|---|---|---|---|
| ||||