Pinaceae
| Pinaceae | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Jurássico–recente | |||||||||||||
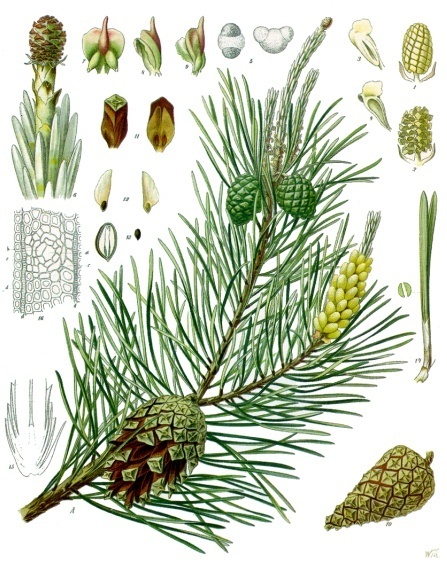 Representação das principais características de Pinaceae em Pinus sylvestris (imagem do Köhlers Medizinal Pflanzen). | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Subfamílias e géneros | |||||||||||||
| |||||||||||||
| Sinónimos | |||||||||||||
| |||||||||||||





Pinaceae é uma família de coníferas da ordem monotípica Pinales, agrupando cerca de 250 espécies de árvores e arbustos repartidas por 11 géneros[1] e duas subfamílias. A família constitui o maior agrupamento taxonómico entre as Acrogymnospermae, as gimnospermas extantes, e o segundo maior (depois de Cupressaceae) em extensão geográfica da sua área de distribuição natural, presente na maior parte do Hemisfério Norte, com a maioria das espécies em climas temperados, mas variando do subártico ao tropical, formando frequentemente o componente dominante das florestas boreais, costeiras e de montanha. Uma espécie, Pinus merkusii, cresce a sul do equador, no Sueste Asiático.[2] Os principais centros de diversidade encontram-se nas montanhas do sudoeste da China, México, centro do Japão e Califórnia. O grupo inclui múltiplas espécies com interesse económico na produção de madeiras, fibra de celulose e resinas, como os pinheiros, abetos e tsugas, e com usos ornamentais.
Descrição
[editar | editar código-fonte]Com o seu monofiletismo suportado pelas características dos plastídeos de tipo proteico das suas células crivosas, padrão de proembriogenia e ausência de bioflavonoides, a família Pinaceae pertence ao grupo das coníferas (Pinophyta), constituindo o maior agrupamento taxonómico entre as acrogimnospermas, as gimnospermas viventes. Presente no registo fóssil conhecido desde o início do Carbonífero, há 300 milhões de anos, a famíia inclui, na sua presente circunscrição taxonómica, entre 220 e 250 espécies (dependendo da opinião taxonómica) repartidas pelos seguintes géneros: Abies, Cedrus, Keteleeria, Larix, Pseudotsuga, Pseudolarix, Cathaya, Nothotsuga, Picea, Pinus e Tsuga.
Características gerais
[editar | editar código-fonte]Para além de ser a família com maior diversidade, as Pinaceae são o grupo mais importante dentro das coníferas em termos ecológicos e económicos, produzindo madeiras utilizadas nas mais diversas indústrias de construção e fornecendo uma variedade de resinas, incluindo resinas utilizadas na indústria farmacêutica.
O grupo tem ocorrência natural no Hemisfério Norte, com apenas uma espécie, Pinus merkusii, nativa a sul do equador, no Sueste Asiático.[2] Apesar disso, diversas espécies são cultivadas com sucesso no hemisfério sul, sendo Pinus e Cedrus os géneros mais utilizados.
Incluída na ordem Pinales, anteriormente conhecida como Coniferales, as Pinaceae são árvores ou arbustos lenhosos, incluindo muitas das coníferas bem conhecidas pela sua importância comercial, tais como cedros, abetos, tsugas, piñons, lariços, pinheiros e abetos.
Morfologia
[editar | editar código-fonte]Os membros da família Pinaceae são árvoress (raramente arbustoss) que crescem de 2 a 100 m de altura, na sua maioria sempre-verdes (exceto as caducifólias Larix e Pseudolarix), resinosas, com ramos subopostos ou espiralados, e folhas lineares (em forma de agulha) dispostas em espiral.[1] Os embriões de Pinaceae têm de 3 a 24 cotilédones.
O ritidoma e as folhas são geralmente aromáticos. Os ramos são opostos ou verticilados e as folhas podem ser do tipo escamiformes ou aciculiformes que, em Pinus, surgem de 2 a 5 acículas em cada fascículo, e são presas a ramos laterais curtos.[3]
Os membros da família Pinaceae são monóicos, apresentando estruturas reprodutivas denominadas cones que tem como funções principais facilitar a dispersão das sementes. O cone é um eixo reprodutivo com escamas e brácteas organizadas numa filotaxia espiralada. As folhas são modificadas em brácteas e podem ser maiores ou menores que as escamas, que são um ramo fértil reduzido, onde os óvulos se encontram na superfície adaxial nos cones femininos, e os microsporângios na superfície abaxial nos cones masculinos.
O cone masculino (microstróbilo) é pequeno, alongado e disposto em cacho, e o cone feminino (megastróbilo) possui uma morfologia cilíndrica e com um aspecto mais esférico, com escamas lenhosas persistentes e com o ápice espesso, contendo 2 sementes aladas que amadurecem em um período de 2 a 3 anos.[4]
Os cones femininos das coníferas são grandes e geralmente lenhosos, com 2 a 60 cm de comprimento, com numerosas escamas dispostas em espiral e duas sementes aladas em cada escama. Os cones masculinos são pequenos, 0,5 a 6 cm de comprimento, e caem logo após a polinização. A dispersão do pólen é feita pelo vento (anemofilia). A dispersão das sementes é também maioritariamente feita pelo vento (anemocoria), mas algumas espécies têm sementes grandes com asas reduzidas que são dispersas por aves (ornitocoria).
A análise dos cones de Pinaceae revela como a pressão selectiva moldou a evolução do tamanho e função variável dos cones em toda a família. A variação do tamanho dos cones na família resultou provavelmente da variação dos mecanismos de dispersão de sementes disponíveis nos seus ambientes ao longo do tempo. Todas as Pinaceae com sementes de peso inferior a 90 miligramas estão aparentemente adaptadas à dispersão pelo vento. Os pinheiros com sementes maiores que 100 mg são mais susceptíveis de terem beneficiado de adaptações que promovem a dispersão por animais, particularmente por aves.[5] As Pináceas que persistem em zonas onde os esquilos são abundantes não parecem ter desenvolvido adaptações para a dispersão por aves.
As coníferas boreais apresentam muitas adaptações para o inverno. A forma cónica estreita das coníferas do Norte e os seus membros inclinados para baixo ajudam-nas a afastar a neve, e muitas delas alteram sazonalmente a sua bioquímica para se tornarem mais resistentes ao congelamento, o que se designa por "endurecimento" ("hardening").
Filogenia e classificação
[editar | editar código-fonte]História evolucionária
[editar | editar código-fonte]Estima-se que a Pinaceae tenha divergido de outros grupos de coníferas durante o final do Carbonífero, há cerca de 313 milhões de anos.[6] O género extinto de coníferas Schizolepidopsis provavelmente representa membros do grupo troncal das Pinaceae, cujos primeiros bons registos se encontram no período médio-tardio do Triásico, com registos abundantes durante o Jurássico em toda a Eurásia.[7][8]
O membro mais antigo do grupo coroa (descendente do último antepassado comum de todas as espécies vivas) de Pinaceae é o cone Eathiestrobus, conhecido do Jurássico Superior (Kimmeridgiano inferior, 157,3-154,7 milhões de anos atrás) da Escócia,[9] que provavelmente pertence ao grupo pinoide da família.[10][8]
As Pinaceae irradiaram rapidamente durante o Cretáceo Inferior.[6] Os membros dos géneros modernos Pinus (pinheiros), Picea (abeto) e Cedrus (cedro) apareceram pela primeira vez durante o Cretáceo Inferior.[11][12][13] Os géneros extintos do Cretáceo Pseudoaraucaria e Obirastrobus parecem ser membros de Abietoideae, enquanto Pityostrobus parece não ser monofilético, contendo muitos membros díspares de Pinaceae.[10]
Filogenia
[editar | editar código-fonte]Na evolução das coníferas, a análise de dados morfológicos como a inversão dos óvulos e a presença de asa terminal na semente derivadas do material da escama inferem que Pinaceae é um grupo monofilético, considerado grupo irmão das demais coníferas agrupadas na antiga divisão Cupressophyta (que incluía as famílias Araucariaceae, Podocarpaceae, Taxaceae, Cupressaceae e Sciadopityaceae). Tais inferências são corroboradas por análises moleculares em genes dos plastídeos das coníferas.
A utilização das técnicas da filogenética molecular permitiram clarificar as relações filogenéticas no interior do agrupamento taxonómico, aconselhando à redução do número de subfamílias a apenas duas (Pinoideae e Abietoideae), de forma a obter agrupamentos monofiléticos. A partir desses estudos chegou-se aos seguintes cladogramas:
| Ran et al. 2018[14] & Leslie et al. 2018[15][16] | Stull et al. 2021[17][18] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Múltiplos estudos moleculares indicam que, em contraste com as classificações anteriores que a colocavam fora das coníferas, Gnetophyta pode de facto ser o grupo irmão das Pinaceae, com ambas as linhagens a divergirem durante o início e meados do Carbonífero. Esta hipótese é conhecida como a hipótese "gnepina" (ou "GnePin").[19][20] Contudo, uma filogenia revista de 2018 coloca Cathaya como grupo irmão de Pinaceae e não na subfamília Laricoidae com Larix e Pseudotsuga.
Classificação
[editar | editar código-fonte]A classificação das subfamílias e géneros de Pinaceae tem sido objeto de debate no passado. A ecologia, morfologia e história de Pinaceae foram utilizadas como base para métodos de análise da família. Uma publicação de 1891 dividiu a família em duas subfamílias, utilizando o número e a posição dos canais resiníferos na região vascular primária da raiz axial jovem como principal fator de consideração. Numa publicação de 1910, a família foi dividida em duas tribos com base na ocorrência e no tipo de dimorfismo longo-curto dos rebentos.
Posteriormente, com base em critérios essencialmente morfológicos, a família Pinaceae foi tradicionalmente dividida subfamílias e géneros com base na consideração das características da anatomia do cone ovulado entre os membros extantes e fósseis da família. Dentro da família Pinaceae, as análises baseadas em caracteres de anatomia microscópica e morfologia de cones, pólen, sementes e folhas levaram à criação de quatro subfamílias: Pinoideae (Pinus), Piceoideae (Picea), Laricoideae (Larix, Cathaya e Pseudotsuga) e Abietoideae (Abies, Cedrus, Pseudolarix, Keteleeria, Northotsuga e Tsuga).[21] Assim, nessa classificação tradicional, os 11 géneros estão agrupados em quatro subfamílias, com base na anatomia microscópica e na morfologia dos cones, pólen, madeira, sementes e folhas:[22]
- Subfamília Pinoideae (monotípica, género-tipo Pinus): os cones são bienais, raramente trienais, com o crescimento das escamas de cada ano distinto, formando um umbo em cada escama, a base da escama do cone é larga, ocultando totalmente as sementes da vista abaxial (abaixo dos vasos do floema), a semente não tem vesículas de resina, a asa da semente segura a semente num par de garras, as folhas têm bandas estomáticas primárias adaxiais (acima do xilema) ou igualmente em ambas as superfícies.
- Subfamília Piceoideae (monotípica, género-tipo Picea): os cones são anuais, sem umbo distinto, a base da escama do cone é larga, ocultando totalmente as sementes da visão abaxial, a semente não tem vesículas de resina, é enegrecida, a asa da semente segura a semente frouxamente num cálice, as folhas têm bandas estomáticas primárias adaxiais (acima do xilema) ou igualmente em ambas as superfícies.
- Subfamília Laricoideae (género-tipo Larix, Pseudotsuga, e Cathaya): os cones são anuais, sem umbo distinto, a base da escama do cone é larga, ocultando totalmente as sementes da visão abaxial, a semente não tem vesículas de resina, é esbranquiçada, a asa da semente segura a semente firmemente num cálice, as folhas têm bandas estomáticas primárias apenas abaxiais.
- Subfamília Abietoideae (género-tipo Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga e Tsuga): os cones são anuais, sem um umbo distinto, a base da escama do cone é estreita, com as sementes parcialmente visíveis em vista abaxial, a semente tem vesículas de resina, a asa da semente mantém a semente firmemente num cálice, as folhas têm bandas estomáticas primárias apenas na face abaxial.
Porém, análises moleculares posteriores mostraram que o género Cathaya está mais próximo do género Picea, e o género Cedrus está mais próximo de ser o grupo irmão dos demais géneros.[23]
Considerando a árvore filogenética construída para o agrupamento Pinales, na sua presente circunscrição taxonómica obtém-se a seguinte classificação para Pinales e Pinaceae:[24]
- Pinales
- Família Pinaceae
- Subfamília Abietoideae
- Subfamília Pinoideae
Distribuição geográfica e ocorrência no Brasil
[editar | editar código-fonte]Pinaceae são encontradas em ampla distribuição principalmente no Hemisfério Norte, ocupando desde regiões temperadas até o Círculo Polar Ártico, e desde solos bem drenados até os permanentemente saturados de água.[3]
Podem ser encontradas também no Hemisfério Sul, mas não são espécies nativas.[25] No Brasil, as sete espécies estão distribuídas no Centro-Oeste (Distrito-Federal e Goias), no Sudeste (Espirito Santo, Minas Gerais, Rio de Janeiro e São Paulo) e no Sul do país (Paraná, Rio Grande do Sul e Santa Catarina). Os principais domínios fitogeográficos são Cerrado e Mata Atlântica.[26]
Assim, apesar de nenhuma das espécies de Pinaceae existentes no Brasil ser espécie nativa, ou seja, são na totalidade espécies introduzidas no território brasileiro, nalguns casos estão naturalizadas (género Pinus):[26]
Pinaceae Spreng. ex F.Rudolphi
- Pinus caribaea Morelet
- Pinus clausa (Chapm. ex Engelm.) Sarg.
- Pinus elliottiL.
- Pinus glabra Walter
- Pinus oocarpa Schiede ex Schltdl.
- Pinus pseudostrobus Lindl.
- Pinus taeda L.
Ecologia
[editar | editar código-fonte]As pressões externas sobre as plantas têm a capacidade de alterar a estrutura e a composição dos ecossistemas florestais. O stress externo mais comum que as Pinaceae sofrem é o ataque de herbívoros e agentes patogénicos, que muitas vezes leva à morte da árvore.[27] Para combater estas pressões, as árvores precisam de se adaptar ou desenvolver defesas contra os seus efeitos. As Pinaceae desenvolveram uma miríade de defesas mecânicas e químicas, ou uma combinação das duas, para se protegerem contra os antagonistas.[28]
As Pinaceae têm a capacidade de regular uma combinação de estratégias mecânicas e estratégias químicas para promover as suas defesas.[29] As defesas das Pinaceae são predominantes na casca das árvores. Esta parte da árvore contribui com uma complexa barreira defensiva contra antagonistas externos[30] e por isso as defesas constitutivas e as defesas induzidas encontram-se ambas na casca.[30][31][32]
As defesas constitutivas são tipicamente a primeira linha de defesa usadas contra antagonistas e podem incluir células esclerificadas, células lignificadas da periderme e compostos secundários como compostos fenólicos e resinas.[33][30][31] As defesas constitutivas são sempre expressas e oferecem proteção imediata contra os invasores, mas também podem ser derrotadas por antagonistas que desenvolveram adaptações a estes mecanismos de defesa.[33][30] Um dos compostos secundários comuns utilizados pelas Pinaceae são os compostos fenólicos ou polifenóis. Estes compostos secundários são conservados nos vacúolos das células polifenólicas do parênquima (PP) no floema secundário.[34][32]
As respostas com base em defesas induzidas precisa ser activada por determinados sinais, tais como danos causados por herbívoros ou outros sinais bióticos.[33] Um mecanismo de defesa induzido comum utilizado pelas "Pinaceae" é a produção de resinas.[35] As resinas são assim uma das principais defesas utilizadas contra ataques por herbívoros.[28] As resinas são defesas de curto prazo que são compostas por uma combinação complexa de monoterpenos (C10) e sesquiterpenos (C15) voláteis e ácidos resínicos diterpenos não voláteis (C20).[28][35]
Estes compostos são produzidos e armazenados em áreas secretoras especializadas conhecidas como dutos de resina, bolhas de resina ou cavidades de resina.[35] As resinas têm a capacidade de lavar, prender, afastar antagonistas e também estão envolvidas na vedação de feridas.[34] São um mecanismo de defesa eficaz porque têm efeitos tóxicos e inibitórios sobre invasores, como insectos ou agentes patogénicos.[36]
As resinas podem ter-se desenvolvido como uma defesa evolutiva contra os ataques do carunchos da casca, um grupo de insetos que inclui Cerambycidae e a subfamília Scolytinae, que causa graves danos em florestas.[35] Uma resina bem conhecida presente nas Pinaceae é a oleorresina. Descobriu-se que a oleorresina é uma parte valiosa do mecanismo de defesa das coníferas contra o ataques bióticos.[36] Estes compostos são encontradas em tecidos secretores nos caules, raízes e folhas das árvores.[36] A oleorresina também é usada como base para classificar as coníferas.[36]
O tópico dos mecanismos de defesa na família Pinaceae é uma área de estudo muito ativa, com numerosos estudos em curso. Muitos destes estudos utilizam o metil jasmonato (MJ) como antagonista.[31][32][37] Sabe-se que o jasmonato de metilo é capaz de induzir respostas de defesa nos caules de múltiplas espécies de Pinaceae.[31][37] Verificou-se que o MJ estimula a ativação das células PP e a formação de ductos resiníferos traumáticos no xilema (TD). Estas são estruturas que estão envolvidas na libertação de compostos fenólicos e resinas, ambos mecanismo de defesa.[31][32]
Importância económica
[editar | editar código-fonte]Muitos animais, como aves e esquilos por exemplo, obtêm alimento a partir das sementes do pinheiro, as quais foram importantes na alimentação de alguns povos indígenas do Oeste dos Estados Unidos.[3] As sementes de algumas espécies de Pinus são consumidas (os pinhões),[38][39] mantendo importante valor económico.
As espécies de Pinaceae fornecem a maior parte da madeira macia utilizada no mundo. A sua madeira é utilizada na construção naval. Fornecem as matérias-primas para a pasta de papel e para as indústrias química e farmacêutica (óleos essenciais). A madeira de árvores como Pinus, Pseudotsuga, Picea, Tsuga, Larix, Abies e Cedrus são mundialmente valorizadas para a construção, e as fibras longas do Pinus são utilizadas como celulose para a fabricação de vários tipos de papéis.[4]
Muitas espécies e os seus cultivares são usados como planta ornamental em parques e jardins, especialmente nas zonas temperadas. Muitas espécies são importantes árvores florestais. São plantadas como quebra-ventos, como controlo de erosão (montanhas) e em medidas de reflorestação.[38]
Referências
[editar | editar código-fonte]- ↑ a b Aljos Farjon (1998). World Checklist and Bibliography of Conifers. [S.l.]: Royal Botanic Gardens, Kew. ISBN 978-1-900347-54-9
- ↑ a b http://www.conifers.org/pi/Pinus_merkusii.php
- ↑ a b c Judd; et al. (2009). Plant systematics: a phylogenetic approach (em inglês). [S.l.]: Sunderland. pp. 172–173
- ↑ a b Reyes, Andrés E.L. «Gimnospermas». www.esalq.usp.br. Consultado em 13 de dezembro de 2017
- ↑ Craig W. Benkman (1995). «Wind dispersal capacity of pine seeds and the evolution of different seed dispersal modes in pines» (PDF). Oikos. 73 (2): 221-224. JSTOR 3545911. doi:10.2307/3545911
- ↑ a b Leslie, Andrew B.; Beaulieu, Jeremy; Holman, Garth; Campbell, Christopher S.; Mei, Wenbin; Raubeson, Linda R.; Mathews, Sarah (2018). «An overview of extant conifer evolution from the perspective of the fossil record». American Journal of Botany (em inglês). 105 (9): 1531–1544. ISSN 1537-2197. PMID 30157290. doi:10.1002/ajb2.1143
- ↑ Domogatskaya, Ksenia V.; Herman, Alexei B. (maio de 2019). «New species of the genus Schizolepidopsis (conifers) from the Albian of the Russian high Arctic and geological history of the genus». Cretaceous Research (em inglês). 97: 73–93. doi:10.1016/j.cretres.2019.01.012
- ↑ a b Matsunaga, Kelly K. S.; Herendeen, Patrick S.; Herrera, Fabiany; Ichinnorov, Niiden; Crane, Peter R.; Shi, Gongle (10 de maio de 2021). «Ovulate Cones of Schizolepidopsis ediae sp. nov. Provide Insights into the Evolution of Pinaceae». International Journal of Plant Sciences. 182 (6): 490–507. ISSN 1058-5893. doi:10.1086/714281
- ↑ Rothwell, Gar W.; Mapes, Gene; Stockey, Ruth A.; Hilton, Jason (Abril de 2012). «The seed cone Eathiestrobus gen. nov.: Fossil evidence for a Jurassic origin of Pinaceae». American Journal of Botany (em inglês). 99 (4): 708–720. PMID 22491001. doi:10.3732/ajb.1100595
- ↑ a b Smith, Selena Y.; Stockey, Ruth A.; Rothwell, Gar W.; Little, Stefan A. (2 de janeiro de 2017). «A new species of Pityostrobus (Pinaceae) from the Cretaceous of California: moving towards understanding the Cretaceous radiation of Pinaceae». Journal of Systematic Palaeontology (em inglês). 15 (1): 69–81. ISSN 1477-2019. doi:10.1080/14772019.2016.1143885
- ↑ Blokhina, N. I.; Afonin, M. (2007). «Fossil wood Cedrus penzhinaensis sp. nov. (Pinaceae) from the Lower Cretaceous of north-western Kamchatka (Russia)». Acta Paleobotanica (em inglês). 47: 379–389. Consultado em 29 de maio de 2021
- ↑ Ashley A. Klymiuk; Ruth A. Stockey (2012). «A Lower Cretaceous (Valanginian) seed cone provides the earliest fossil record for Picea (Pinaceae)». American Journal of Botany. 99 (6): 1069–1082. PMID 22623610. doi:10.3732/ajb.1100568
- ↑ Patricia E. Ryberg; Gar W. Rothwell; Ruth A. Stockey; Jason Hilton; Gene Mapes; James B. Riding (2012). «Reconsidering Relationships among Stem and Crown Group Pinaceae: Oldest Record of the Genus Pinus from the Early Cretaceous of Yorkshire, United Kingdom». International Journal of Plant Sciences. 173 (8): 917–932. doi:10.1086/667228
- ↑ Ran, Jin-Hua; Shen, Ting-Ting; Wu, Hui; Gong, Xun; Wang, Xiao-Quan (1 de dezembro de 2018). «Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis». Molecular Phylogenetics and Evolution (em inglês). 129: 106–116. ISSN 1055-7903. PMID 30153503. doi:10.1016/j.ympev.2018.08.011
- ↑ Leslie, Andrew B.; Beaulieu, Jeremy; Holman, Garth; Campbell, Christopher S.; Mei, Wenbin; Raubeson, Linda R.; Mathews, Sarah; et al. (2018). «An overview of extant conifer evolution from the perspective of the fossil record». American Journal of Botany. 105 (9): 1531-1544. doi:10.1002/ajb2.1143
- ↑ Leslie, Andrew B.; et al. (2018). «ajb21143-sup-0004-AppendixS4» (PDF)
- ↑ Stull, Gregory W.; Qu, Xiao-Jian; Parins-Fukuchi, Caroline; Yang, Ying-Ying; Yang, Jun-Bo; Yang, Zhi-Yun; Hu, Yi; Ma, Hong; Soltis, Pamela S.; Soltis, Douglas E.; Li, De-Zhu; Smith, Stephen A.; Yi, Ting-Shuang; et al. (2021). «Gene duplications and phylogenomic conflict underlie major pulses of phenotypic evolution in gymnosperms». Nature Plants. 7 (8): 1015–1025. PMID 34282286. doi:10.1038/s41477-021-00964-4
- ↑ Stull, Gregory W.; et al. (2021). «main.dated.supermatrix.tree.T9.tre». Figshare. doi:10.6084/m9.figshare.14547354.v1
- ↑ Stull, Gregory W.; Qu, Xiao-Jian; Parins-Fukuchi, Caroline; Yang, Ying-Ying; Yang, Jun-Bo; Yang, Zhi-Yun; Hu, Yi; Ma, Hong; Soltis, Pamela S.; Soltis, Douglas E.; Li, De-Zhu (19 de Julho de 2021). «Gene duplications and phylogenomic conflict underlie major pulses of phenotypic evolution in gymnosperms». Nature Plants (em inglês). 7 (8): 1015–1025. ISSN 2055-0278. PMID 34282286. doi:10.1038/s41477-021-00964-4
- ↑ Ran, Jin-Hua; Shen, Ting-Ting; Wang, Ming-Ming; Wang, Xiao-Quan (2018). «Phylogenomics resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and angiosperms». Proceedings of the Royal Society B: Biological Sciences. 285 (1881). 20181012 páginas. PMC 6030518
. PMID 29925623. doi:10.1098/rspb.2018.1012
- ↑ Price, Robert A.; Olsen-Stojkovich, Jeanine; Lowenstein, Jerold M. (1987). «Relationships Among the Genera of Pinaceae: An Immunological Comparison». Systematic Botany (em inglês). 12 (1): 91–97. doi:10.2307/2419217
- ↑ Robert A. Price, Jeanine Olsen-Stojkovich & Jerold M. Lowenstein (1987). «Relationships among the genera of Pinaceae: an immunological comparison». Systematic Botany. 12 (1): 91–97. JSTOR 2419217. doi:10.2307/2419217
- ↑ Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as refs de nome:3a - ↑ APweb: Pinales.
- ↑ Simpson, M.G. (2010). Plant Systematics. Amsterdam: Elsevier. pp. 148–151
- ↑ a b «Pinaceae in Flora do Brasil 2020 em construção». Jardim Botânico do Rio de Janeiro. Consultado em 2 de novembro de 2017
- ↑ Cherubini, Paolo; Fontana, Giovanni; Rigling, Daniel; Dobbertin, Matthias; Brang, Peter; Innes, John L. (2002). «Tree-Life History Prior to Death: Two Fungal Root Pathogens Affect Tree-Ring Growth Differently». Journal of Ecology. 90 (5): 839–850. JSTOR 3072253. doi:10.1046/j.1365-2745.2002.00715.x
- ↑ a b c Zulak, K. G.; Bohlmann, J. (2010). «Terpenoid biosynthesis and specialized vascular cells of conifer defense. - Semantic Scholar». Journal of Integrative Plant Biology. 52 (1): 86–97. PMID 20074143. doi:10.1111/j.1744-7909.2010.00910.x
- ↑ Franceschi, Vincent R.; Krokene, Paal; Christiansen, Erik; Krekling, Trygve (1 de agosto de 2005). «Anatomical and chemical defenses of conifer bark against bark beetles and other pests». New Phytologist (em inglês). 167 (2): 353–376. ISSN 1469-8137. PMID 15998390. doi:10.1111/j.1469-8137.2005.01436.x
- ↑ a b c d Franceschi, V. R., P. Krokene, T. Krekling, and E. Christiansen. 2000. Phloem parenchyma cells are involved in local and distance defense response to fungal inoculation or bark-beetle attack in Norway spruce (Pinaceae). American Journal of Botany 87:314-326.
- ↑ a b c d e Hudgins, J. W.; Christiansen, E.; Franceschi, V. R. (1 de março de 2004). «Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: a phylogenetic perspective». Tree Physiology (em inglês). 24 (3): 251–264. ISSN 0829-318X. PMID 14704135. doi:10.1093/treephys/24.3.251
- ↑ a b c d Krokene, P.; Nagy, N. E.; Solheim, H. (1 de janeiro de 2008). «Methyl jasmonate and oxalic acid treatment of Norway spruce: anatomically based defense responses and increased resistance against fungal infection». Tree Physiology (em inglês). 28 (1): 29–35. ISSN 0829-318X. PMID 17938111. doi:10.1093/treephys/28.1.29
- ↑ a b c Sampedro, L. (1 de setembro de 2014). «Physiological trade-offs in the complexity of pine tree defensive chemistry». Tree Physiology (em inglês). 34 (9): 915–918. ISSN 0829-318X. PMID 25261122. doi:10.1093/treephys/tpu082
- ↑ a b Nagy, N. E.; Krokene, P.; Solheim, H. (1 de fevereiro de 2006). «Anatomical-based defense responses of Scots pine (Pinus sylvestris) stems to two fungal pathogens». Tree Physiology (em inglês). 26 (2): 159–167. ISSN 0829-318X. PMID 16356912. doi:10.1093/treephys/26.2.159
- ↑ a b c d Nagy, Nina E.; Franceschi, Vincent R.; Solheim, Halvor; Krekling, Trygve; Christiansen, Erik (1 de março de 2000). «Wound-induced traumatic resin duct development in stems of Norway spruce (Pinaceae): anatomy and cytochemical traits». American Journal of Botany (em inglês). 87 (3): 302–313. ISSN 1537-2197. JSTOR 2656626. PMID 10718991. doi:10.2307/2656626
- ↑ a b c d Lewinsohn, Efraim; Gijzen, Mark; Croteau, Rodney (1 de maio de 1991). «Defense Mechanisms of Conifers: Differences in Constitutive and Wound-Induced Monoterpene Biosynthesis Among Species». Plant Physiology (em inglês). 96 (1): 44–49. ISSN 0032-0889. PMC 1080711
- ↑ a b Fäldt, Jenny; Martin, Diane; Miller, Barbara; Rawat, Suman; Bohlmann, Jörg (1 de janeiro de 2003). «Traumatic resin defense in Norway spruce (Picea abies): Methyl jasmonate-induced terpene synthase gene expression, and cDNA cloning and functional characterization of (+)-3-carene synthase». Plant Molecular Biology (em inglês). 51 (1): 119–133. ISSN 0167-4412. PMID 12602896. doi:10.1023/A:1020714403780
- ↑ a b Christopher J. Earle, Michael P. Frankis: Pinaceae in The Gymnosperm Database, 2019. (inglês.)
- ↑ Pinaceae bei Plants For A Future
Galeria
[editar | editar código-fonte]-
Cone de Pinus attenuata
-
-
-
-
-
-
-
-
-
-
Larix lyallii (no outono)











Ligações externas
[editar | editar código-fonte]- Arboretum de Villardebelle French Arboretum of conifers around the world
- Gymnosperm Database – Pinaceae
- Pinaceae on the web page of the Tree-of-Life project
- 40 Pine Trees From Around the World by The Spruce
- Predefinição:Jepson eFlora, covers Californian species and much of western North America
- Pinaceae in Flora of North America
- Pinus in USDA Plants Database