Proteína transmembrana
Unha proteína transmembrana (PT) é unha proteína que atravesa unha membrana da célula dun lado a outro asomando polos dous lados da membrana. Moitas destas proteínas funcionan como portas de entrada ou "peiraos de carga" que permiten o transporte de substancias específicas (ou impiden o doutras) a través das membranas celulares, as cales entrarán na célula ou sairán (caso, por exemplo, de produtos residuais). Como resposta ao recoñecemento da forma de certas moléculas que se unen a elas, estas proteínas poden adoptar un modo especial de pregamento que leve a substancia que se vai transportar ao outro lado da membrana.
As proteínas transmembrana agréganse e precipitan en auga. Para extraelas da membrana requírense deterxentes ou solventes non polares, aínda que algúns deles (os barrís beta) poden extraerse utilizando axentes desnaturalizantes.
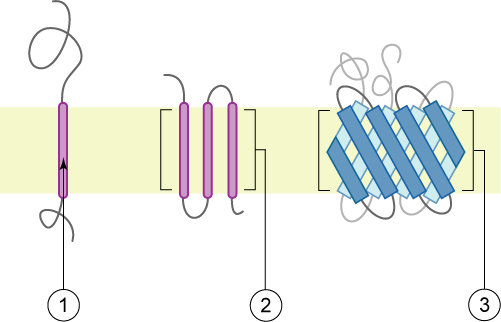
1. proteína transmembrana en hélice α simple (proteína de membrana bitópica)
2. proteína transmembrana en hélice α politópica
3. proteína transmembrana en folla β politópica
A membrana representouse en castaño claro.
Todas as proteínas transmembrana son proteínas integrais de membrana (incluídas na membrana), pero non todas as proteínas integrais de membrana son proteínas transmembrana, xa que moitas non chegan ao outro lado da membrana.[1]
Tipos
editarClasificación estrutural
editarHai dous tipos básicos de proteínas transmembrana:[2]
- En hélice alfa. Estas proteínas están presentes nas membranas internas de células bacterianas gramnegativas ou na membrana plasmática dos eucariotas, e ás veces na membrana bacteriana externa.[3] Esta é a principal categoría de proteínas transmembrana. Nos humanos estímase que o 27% de todas as proteínas son proteínas de membrana en hélice alfa.[4]
- En barril beta. Estas proteínas encontráronse só ata agora en membranas externas de bacterias gramnegativas, paredes celulares de bacterias grampositivas, e na membrana externa das mitocondrias e cloroplastos. Todas as proteínas transmembrana en barril beta teñen unha topoloxía sándwich (up-and-down) moi simple, que pode reflectir a súa orixe común evolutiva, e un mecanismo similar de pregado.
Clasificación topolóxica
editarEsta clasificación refírese á posición dos dominios N- e C-terminal. Os tipos I, II, e III son moléculas dun só paso da cadea polipeptídica a través da membrana, mentres que o tipo IV son moléculas de varios pasos. As proteínas transmembrana de tipo I están ancoradas á membrana lipídica cunha secuencia áncora e os seus dominios N-terminais están dirixidos ao lume do retículo endoplasmático (RE) durante a síntese (e ao espazo extracelular, se as proteínas maduras están localizadas no plasmalema). As de tipo II e III están ancoradas cunha secuencia áncora sinal, e as de tipo II están destinadas ao lume do RE polo seu dominio C-terminal, mentres que nas de tipo III é o seu dominio N-terminal o que as destina ao lume do RE. O tipo IV subdivídese en IV-A, co dominio N-terminal destinado ao citosol, e o IV-B, co dominio N-terminal destinado ao lume.[5] As implicacións para a división nos catro tipos póñense de manifesto especialmente no momento da translocación e a tradución de proteínas en ribosomas unidos ao RE rugoso, cando as proteínas teñen que pasar a través da membrana do RE nunha dirección que depende do tipo.
Estabilidade termodinámica e pregamento
editarEstabilidade das proteínas transmembrana en hélice α
editarAs proteínas transmembrana en hélice alfa son infrecuentemente estables segundo os estudos de desnaturalización térmica realizados, porque non se despregan totalmente dentro das membranas (o despregamento completo requiriría a rotura do moitos enlaces de hidróxeno da hélice α no medio non polar). Por outra parte, estas proteínas préganse incorrectamente con facilidade, debido á agregación non nativa nas membranas, transición aos estados de glóbulo fundido, formación de pontes disulfuro non nativas, ou o despregamento de rexións periféricas e bucles non regulares que son localmente menos estables
Tamén é importante definir exactamente o que é o estado despregado. O estado despregado de proteínas de membrana en micelas deterxentes é diferente do dos experimentos de desnaturalización térmica. Este estado representa unha combinación de hélices α hidrofóbicas pregadas e segmentos parcialmente despregados cubertos polo deterxente. Por exemplo, a bacteriorrodopsina "despregada" en micelas de SDS ten catro hélices α transmembrana pregadas, mentres que o resto da proteína está situada na interface micela-auga e pode adoptar diferentes tipos de estruturas anfifílicas non nativas. As diferenzas de enerxía libre entre eses estados desnaturalizados por deterxentes e os nativos son similares para estabilidades de proteínas hidrosolubles (< 10 kcal/mol).
Pregamento de proteínas transmembrana en hélice α
editarO repregamento de proteínas transmembrana en hélice α in vitro é tecnicamente difícil. Hai relativamente poucos exemplos de experimentos que conseguisen renaturalizacións con éxito, como o da bacteriorrodopsina. In vivo todas esas proteínas están normalmente pregadas co-traducionalmente no grande translocón transmembrana. A canle translocón proporciona un ambiente moi heteroxéneo para as hélices α transmembrana nacentes. Unha hélice α anfifílica relativamente polar pode adoptar unha orientación transmembrana no translocón (aínda que estaría na superficie da membrana ou despregada in vitro), porque os seus residuos polares poden orientarse cara á canle central chea de auga do translocón. Este mecanismo é necesario para a incorporación de hélices α polares en estruturas de proteínas transmembrana. As hélices anfifílicas permanecen unidas ao translocón ata que a proteína é completamente sintetizada e pregada. Se a proteína permanece despregada e unida ao translocón por moito tempo, é degradada por sistemas celulares específicos de "control de calidade".
Estabilidade e pregamento de proteínas transmembrana en barril β
editarA estabilidade de proteínas transmembrana en barril β é similar á estabilidade de proteínas hidrosolubles, segundo os estudos de desnaturalización química. O seu pregamento in vivo está facilitado por chaperonas hidrosolubles, como a proteína Skp [1].
Estruturas 3D
editarTransportadores impulsados pola absorción de luz
editar- Proteínas do tipo da bacteriorrodopsina como a rodopsina (ver tamén opsina)[2]
- Centros de reacción fotosintéticos bacterianos e fotosistemas I e II [3]
- Complexos recolectores de luz de bacterias e cloroplastos [4]
Transportadores impulsados por oxidorreducións
editar- Proteínas do tipo do citocromo b transmembrana [5]: coencima Q - citocromo c redutase (citocromo bc1 ); complexo do citocromo b6f; formato deshidroxenase, nitrato redutase respiratoria; succinato - coencima Q redutase (fumarato redutase); e succinato deshidroxenase. Ver cadea de transporte de electróns.
- Ctocromo c oxidases [6] de bacterias e mitocondrias.
Transportadores impulsados por potenciais electroquímicos
editarTransportadores impulsados pola hidrólise de enlaces fosfato
editar- ATPase de calcio tipo P (cinco conformacións distintas) [8]
- Reguladores da ATPase de calcio fosfolamban e sarcolipina [9]
- Transportadores ABC: BtuCD, transportador multidrogas, e transportador captador de molibdato
- Translocón da vía secretora xeral (Sec) (preproteína translocase SecY) [10]
Transportadores (uniportadores, simportadores, antiportadores)
editar- Proteínas transportadoras (carriers) mitocondriais [11]
- Superfamilia do Facilitador Maior (transportador de glicerol-3-fosfato, lactosa permease, e transportador multidrogas EmrD) [12]
- Resistencia-nodulación-división celular (transportador AcrB de fluxo multidrogas, ver resistencia a multidrogas)[13]
- Simportador dicarboxilato/aminoácido:catión (simportador protón glutamato) [14]
- Antiportador catión monovalente/protón (antiportador sodio/protón 1 NhaA) [15]
- Simportador de sodio neurotransmisor [16]
- Transportadores de amoníaco [17]
- Transportador drogas/metabolito (transportador de resistencia a multidrogas pequeno EmrE - as estruturas que se obtiveron foron retiradas -retractadas- por seren erradas) [18]
Canle en hélice alfa incluíndo as canles iónicas
editar- De tipo canle iónica regulada por voltaxe, como as canles de potasio KcsA e KvAP, e canle de ión potasio rectificadora entrante Kirbac [19]
- Canle mecanosensible de gran condutancia, MscL [20]
- Canle iónica mecanosensible de pequena condutancia (MscS) [21]
- Transportadores de ións metálicos CorA [22]
- Canle iónica regulada por ligando dos receptores de neurotransmisores (receptor da acetilcolina) [23]
- Acuaporinas [24]
- Canles de cloruro [25]
- Proteínas auxiliares da membrana externa (transportadores de polisacáridos) [26] - proteínas transmembrana en hélice α da membrana bacteriana externa gramnegativa.
Encimas
editar- Metano monooxixenase [27]
- Romboide protease [28]
- Proteína de formación de pontes disulfuro (complexo DsbA-DsbB) [29]
Proteínas con áncoras transmembrana en hélice alfa
editar- Dominio de dimerización transmembrana do receptor de células T [30]
- Complexo citocromo c nitrito redutase [31]
- Esteril-sulfato sulfohidrolase [32]
- Estanina [33]
- Dímero de glicoforina A [34]
- Proteína da cuberta maior de inovirus (fago filamentoso) [35]
- Pilina [36]
- Proteína asociada ao surfactante pulmonar [37]
- Monoamino oxidases A e B [38],
- Ácido graxo amida hidrolase [6]
- Citocromo P450 oxidases [39],
- Corticosteroide 11β-deshidroxenases [40].
- Peptidase do péptido sinal [41]
- Protease de membrana específica para un homólogo da estomatina [42]
Barrís β compostos por unha soa cadea polipeptídica
editar- Barrís beta desde oito cadeas beta e con "número de cizalla" de dez (n=8, S=10) [43]. Entre elas están:
- Dominio transmembrana de tipo OmpA (OmpA),
- Familia das proteínas da membrana externa relacionadas coa virulencia (OmpX),
- Familia W de proteínas da membrana externa (OmpW),
- Familia de proteínas de acilación do lípido A e resistencia a péptidos antimicrobianos (PagP)
- Lípido A desacilase PagL, e
- Porinas da familia opacidade (NspA)
- Dominio autortransportador (n=12,S=14') [44]
- Familia da proteína transportadora da membrana externa FadL, incluíndo o transportador de ácidos graxos FadL (n=14,S=14) [45]
- Familia da porina bacteriana xeral, coñecida como porina trimérica (n=16,S=20) [46]
- Maltoporina, ou porinas de azucres (n=18,S=22) [47]
- Porina específica de nucleósidos (n=12,S=16) [48]
- Fosfolipase A1 da membrana externa (n=12,S=16) [49]
- Receptores dependentes de TonB e os seus dominios plug. Son canles da membrana externa regulados por ligando (n=22,S=24), entre os que está o transportador da cobalamina BtuB, o receptor FptA Fe(III)-pioquelina, o receptor FepA, o receptor da captación de hidroxamato férrico FhuA, o transportador FecA, e o receptor de pioverdina FpvA [50]
- Familia da proteína da membrana externa OpcA (n=10,S=12), que inclúe a protease da membrana externa OmpT e a proteína adhesina/invasina OpcA [51]
- Familia de porinas de proteína G da membrana externa (n=14,S=16) [52]
Nota: n e S son, respectivamente, o número de cadeas beta e o "número de cizalla" (shear number)[7] do barril beta.
Barrís β compostos por varias cadeas polipeptídicas
editar- Autotransportador trimérico (n=12,S=12) [53]
- Proteínas de fluxo da membrana externa, tamén chamadas factores da membrana externa triméricos (n=12,S=18), como o TolC e proteínas de resistencia a multidrogas [54]
- MspA porina (octámero, n=S=16) e α-hemolisina (heptámero n=S=14) [55]. Estas proteínas son segregadas.
Ver tamén gramicidina A [56], un péptido que forma unha hélice β transmenbrana dimérica. Tamén se segrega por bacterias grampositivas.
Notas
editar- ↑ Steven R. Goodman (2008). Medical cell biology. Academic Press. pp. 37–. ISBN 978-0-12-370458-0. Consultado o 24 November 2010.
- ↑ Jin Xiong (2006). Essential bioinformatics. Cambridge University Press. pp. 208–. ISBN 978-0-521-84098-9. Consultado o 13 November 2010.
- ↑ Entre as proteínas en hélice alfa de membranas externas bacterianas están e estanina e certas lipoproteínas, e outras
- ↑ Almén MS, Nordström KJ, Fredriksson R, Schiöth HB (2009). "Mapping the human membrane proteome: a majority of the human membrane proteins can be classified according to function and evolutionary origin". BMC Biol. 7: 50. PMC 2739160. PMID 19678920. doi:10.1186/1741-7007-7-50.
- ↑ Harvey Lodish etc.; Molecular Cell Biology, Sixth edition, p.546
- ↑ Bracey MH, Hanson MA, Masuda KR, Stevens RC, Cravatt BF (2002). "Structural adaptations in a membrane enzyme that terminates endocannabinoid signaling". Science 298 (5599): 1793–6. PMID 12459591. doi:10.1126/science.1076535.
- ↑ Murzin AG, Lesk AM, Chothia C (1994). "Principles determining the structure of beta-sheet barrels in proteins. I. A theoretical analysis". J. Mol. Biol. 236 (5): 1369–81. PMID 8126726. doi:10.1016/0022-2836(94)90064-7.
Véxase tamén
editarOutros artigos
editarLigazóns externas
editar- Bruce Alberts et al. Molecular Biology of the Cell. 4th edition. Membrane proteins