As gramíneas ou poáceas (PoaceaeBarnhart) son unha familia de plantasherbáceas, ou moi raramente leñosas, pertencente á ordePoales das monocotiledóneas (Liliopsida). Con máis de 670 xéneros e preto de 10.000 especies descritas, as gramíneas son a cuarta familia con maior riqueza de especies logo das compostas (Asteraceae), as orquídeas (Orchidaceae) e as leguminosas (Fabaceae); mais, definitivamente, é a primeira en importancia económica global.[1] De feito, a maior parte da dieta dos seres humanos provén das gramíneas, tanto de xeito directo (grans de cereais e os seus derivados, como fariñas e aceites) ou indirecto (carne, leite e ovos que proveñen do gando e das aves de curral que se alimentan de pastos ou grans). É unha familia cosmopolita, que conquistou a maioría dos nichos ecolóxicos do planeta, desde as zonas desérticas até os ecosistemas de auga salgada, e desde as zonas deprimidas até os sistemas montañosos máis altos. Esta incomparable capacidade de adaptación está sustentada nunha enorme diversidademorfolóxica, fisiolóxica e reprodutiva e en varias asociacións mutualísticas con outros organismos, que converten ás gramíneas nunha fascinante familia, non só pola súa importancia económica, senón tamén pola súa relevancia biolóxica.
En xeral son herbas, aínda que poden ser leñosas como os bambús tropicais, cespitosas, rizomatosas ou estoloníferas. Pola duración do seu ciclo de vida poden ser anuais, bienais ou perennes. As gramíneas anuais, como é lóxico supor, reprodúcense unha soa vez durante o seu ciclo vital (o caso do trigo ou da avea, por exemplo). As especies perennes, en cambio, poden reproducirse varias veces (unha vez ao ano, normalmente) ou unha soa vez. No primeiro caso as especies denomínanse iteróparas (a maioría das especies de pastos. por exemplo) e no segundo caso, semélparas (as diferentes especies de bambús).
Teñen talos cilíndricos a elípticos na súa sección transversal, articulados, chamados ordinariamente canas, en xeral con nós macizos e entrenós ocos (mais poden ser totalmente macizos como no caso do millo e algúns bambús). Os nós son algo máis grosos que os entrenós e neles nacen as follas e as xemas. Os entrenós son ás veces algo achatados na zona onde se desenvolven as ramificacións. Un pouco máis arriba do nó existe un meristema intercalar en forma de anel que determina o alongamento do talo. Nalgúns xéneros existen de 2 a 6 nós moi próximos entre si (os cales se denominan nós compostos), cada un dos cales leva a súa correspondente folla. En Cynodon dactylon, por exemplo, os nós están en grupos de dous en dous polo que as follas parecen opostas. En xeral os entrenós basais son máis curtos que os superiores; cando hai varios nós basais moi próximos, as follas semellan arrosetadas, é dicir, disponse de xeito tal que simulan formar unha roseta basal de follas. Os principais tipos de talos nas gramíneas son os seguintes:
Talos aéreos:
Talos ascendentes, xeralmente con entrenós curtos na base e gradualmente máis longos cara ao ápice. Poden ser simples ou ramificados.
Talos rastreiros: que medran deitados sobre o chan, arraigando nos nós; frecuentemente presentan nós compostos, como en Stenotaphrum e Cynodon. Cando os talos rastreiros orixinan novas plantas en cada nó denomínanse estolóns.
Talos flotantes, os cales flotan na auga grazas aos entrenós ocos ou á presenza de aerénquima. Talos subterráneos:
Rizomas: existen dous tipos de rizomas nas gramíneas. Un primeiro tipo de rizomas curtos, de crecemento definido, que xeran novas plantas xunto á mata orixinal contribuíndo a aumentar o seu diámetro, por exemplo en Arundo donax e Spartina densiflora. O outro tipo de rizoma é longo, de crecemento indefinido que propaga a planta a certa distancia da mata orixinal. Exemplos deste segundo tipo son Sorghum halepense e Panicum racemosum.
Posúen follas de disposición alterna, dísticas, compostas tipicamente de vaíña, lígula e limbo. A vaíña rodea ao talo, as súas marxes superpóñense mais non se fusionan entre si (só ocasionalmente poden ser atopadas formando un tubo). A lígula é un pequeno apéndice membranoso, ou raramente un grupo de pelos, situado na zona de unión do limbo coa vaíña, na parte adaxial. O limbo (ou lámina) é simple, xeralmente lineal, con nervación paralela. Pode ser aplanado ou ás veces enrolado nun tubo, pode ser continuo coa vaíña ou posuír pecíolo. Ademais desta descrición, é necesario abordar a variabilidade que se pode achar para cada un destes órganos:
Lígula membranosa en avea (Avena sativa).Aurículas en trigo (Triticum aestivum).
Profilo: é a primeira folla de cada innovación, xeralmente está reducida a unha vaíña membranosa, sen lígula nin lámina, que protexe á innovación.
Vaíña: a vaíña nace nun nó e envolve á cana, podendo ser máis curta ou máis larga que o entrenó. Frecuentemente a vaíña fica fendida até a súa base, aínda que os seus bordos superpóñense enrolándose ao redor da cana, pero en moitas especies está parcial ou totalmente ligada polos seus bordos (exemplo, en Bromus) e mesmo a lígula pode formar un anel continuo ao redor da cana, como ocorre en Melica e nalgunhas especies de Poa. As nervaduras da vaíña son numerosas e uniformes, aínda que en especies con canas comprimidas as vaíñas poden presentar unha carena conspicua.
Pseudopecíolo: en moitas bambúseas existe unha contracción entre a lámina e a vaíña que semella un pecíolo; frecuentemente este pseudopecíolo fica articulado coa lámina e é persistente. Tamén hai pseudopecíolos no xénero Pharus e en Setaria palmifolia.
Lámina: xeralmente a lámina é lineal ou lanceolada (forma de lanza), enteira na marxe e con nervaduras paralelas. Nos xéneros tropicais aparecen láminas, como en Pharus e Olyra. A lámina de Neurolepis (Bambusoideae) pode ter até 4 metros de longo. Cando as láminas son anchas e planas, como enZea e en Sorghum, hai unha nervadura central conspicua. En troques nas follas lineais, estreitas, as nervaduras son máis ou menos iguais entre si. Esta característica apréciase perfectamente na primeira prefoliación. Permite, ademais, identificar a certas especies. Por exemplo, as follas conduplicadas son típicas de Stenotaphrum secundatum,Bromus brevis,Dactylis glomerata,Axonopus compressus,Poa lanuginosa, entre outras. As follas convolutas pódense apreciar en Lolium multiflorum,Bromus unioloides,Paspalum dilatatum, entre varios outros exemplos. Nalgúns casos a lámina modifícase facéndose grosa, como en Sporobolus rigens.
Epiderme: os caracteres epidérmicos da folla e das brácteas das gramíneas teñen grande importancia en sistemática e con frecuencia serven para diferenciar determinadas subfamilias ou tribos. En xeral, a disposición das células epidérmicas é diferente nas caras superior e inferior da lámina. Tamén é distinta a disposición sobre os feixes vasculares (chámanse zonas costais) e entre devanditos feixes (zonas intercostales). As células da epiderme das gramíneas pódense agrupar en 5 categorías:
Células silíceas, non posúen cor, recoñécense polo seu brillo particular xa que refractan a luz dun xeito diferente ao resto das células epidérmicas debido a que posúen o lume (o interior da célula) ocupado por sílice. Poden ser redondas, alargadas lonxitudinalmente, en forma de machada de dobre gume, en forma de morna ou contraídas no seu parte central.
Células suberosas, son células curtas, mortas á madurez, coa parede celular suberificada.
Células exodérmicas que comprenden os elementos que sobresaen da superficie da epiderme, os que inclúen: pelos unicelulares, bicelulares ou pluricelulares; pelos capitados, pelos ríxidos e punzantes (aguillóns) máis ou menos silicificados e papilas (células con prominencias cónicas) moi curtas.
Estomas, formados por 2 tipos de células: as células oclusivas ou de peche, en forma de morna, alargadas e engrosadas nos seus extremos, as cales rodean ao orificio por onde se realiza o intercambio gaseoso ou poro estomático, e as células anexas, semilunares, rectangulares ou trapezoidais.
Células longas con paredes lisas e células buliformes sen cor que forman bandas no fondo de fendas sobre a cara superior da lámina. Certos elementos epidérmicos son comúns a todas as Gramíneas, como as células longas, as suberosas, os pelos unicelulares e os aguillóns. Outros son característicos e particulares de certos grupos.
Anatomía foliar: a anatomía da folla das gramíneas ten grande importancia en sistemática. Existen dous tipos de anatomía extremas:
Tipo festucoide, coa vaíña exterior dos feixes vasculares (chamada vaíña parenquimática) pouco diferenciada, con cloroplastos, e a vaíña interior (chamada vaíña mestomática) ben desenvolvida e fortemente engrosada, sen clorofila; ademais o clorénquima non está ordenado en ningunha forma particular. O parénquima sen cor non está presente. Este tipo de anatomía é a típica das Festúceas e certas tribos tropicais como as oríceas.
Tipo panicoide. A vaíña mestomática está apenas desenvolvida ou falta por completo, mentres que a vaíña parenquimática está moi desenvolvida. Neste tipo de anatomía o clorénquima tende a ordenarse en forma radial ao redor dos feixes vasculares, con células lobuladas. O parénquima sen cor está presente. Este tipo anatómico, característico das paníceas, rragrósteas, clorídeas e outras tribos tropicais, denomínase anatomía de Kranz e é propio das gramíneas C4. En contraste, o tipo anatómico festucoide é propio das gramíneas C3. Ademais, existen tipos intermedios por exemplo nas bambúseas, onde tanto a vaíña parenquimática como a mestomática están moi desenvolvidas. As diferenzas na anatomía das follas están asociadas con vías fotosintéticas diferentes. A vía do C3 é máis eficiente en rexións de clima amornado a frío, mentres que a vía do C4 é vantaxosa en rexións de temperaturas altas e baixa humidade do chan. A fisioloxía C3 foi documentada en forma directa en 366 xéneros mentres que a fisioloxía C4 foi documentada en forma directa en 335 xéneros. A fisioloxía intermedia entre C3 e C4 foi observada en Neurachne minor, Steinchisma decipiens (=Panicum decipiens), S. hians e en S. spathellosum.[2][3]
Os macollos ou macollas son a unidade estrutural da maioría das especies de gramíneas. Fórmanse a partir das xemas axilares ou secundarias do meristema basal do eixo principal. Cada un destes brotes secundarios ou macollos comenzan a súa aparición cando as plantas presentan entre dúas e tres follas. Cada un deles, logo de producir as súas primeiras follas, xera o seu propio sistema radicular. A suma ou adición de macollos é o que conforma a estrutura e a forma dunha planta de gramínea. Cando as gramíneas áchanse en estado vexetativo producen continuamente novos macollos e follas. Cada macollo, á súa vez, comezará no seu momento a producir novos macollos.
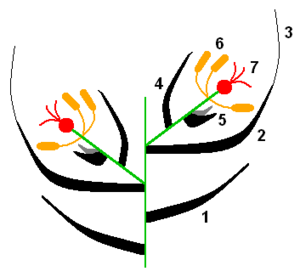
Esquema dunha espigueta: 1. Glumas; 2: Lemma; 3: Aresta; 4: Pálea; 5: Lodículas; 6: Androceo; 7: Xineceo.Detalle dunha espigueta de gramínea, obsérvanse as glumas, a lemma e a pálea.
A inflorescencia elemental das gramíneas é unha pequena espiga formada por unha ou máis flores sentadas sobre un raque articulado, e protexido por brácteas estériles chamadas glumas. Este tipo de inflorescencia recibe o nome de espigueta. As flores poden ser hermafroditas ou unisexuais e presentan un perianto rudimentario de 2 ó 3 pezas, as lodículas ou glumélulas. Estas lodículas son os órganos que, determinan a apertura do antecio ou casa floral durante a floración, permitindo que se expoñan os estigmas e os estames. Os antecios están formados pola lemma, adherida á raquilla, e a pálea inserida sobre o eixo floral que nace sobre a raquilla e soporta aos órganos florais propiamente devanditos. A lemma (ou glumela inferior) ten forma de quilla, e abraza á pálea cos seus bordos. A aresta nace na extremidade da lemma ou no seu dorso. A pálea ou glumela superior é lanceolada, binervada e é como unha tapa que pecha á flor. As glumas están inseridas sobre a raquilla, unha máis abaixo que a outra.
Todos estes elementos son moi variables polo que é conveniente analizalos por separado.
Pedicelo. É o pedúnculo que sostén á espiguilla que pode ser máis ou menos longo ou estar completamente ausente, nese caso as espiguillas son sésiles.
Raquilla, ou eixo da inflorescencia. Adoita levar en cada ángulo un antecio. A raquiña pode estar articulada co pedicelo por baixo das glumas (que entón caen coa espiguilla) ou por encima das glumas (que, entón, son persistentes). A raquiña pode ser tenaz ou fragmentarse á madurez dos froitos.
Glumas. Tipicamente son dúas, alternas, herbáceas, membranosas ou papiráceas, ovadas ou lanceoladas, contraídas lateralmente e máis ou menos carenadas,[4] ou redondeadas, case planas por compresión dorsiventral da espiguilla (Paspalum). Poden levar unha ou varias nervaduras e ser múticas, mucronadas ou aristadas no ápice. Nas oríceas as glumas son rudimentarias ou están ausentes. En cambio, en moitas paníceas hai aparentemente 3 glumas, sendo o superior en realidade unha lemma estéril; ás veces hai unha soa gluma como nos xéneros Monerma e Lolium.
Antecios. Os antecios ('casa floral' en grego) poden ser desde un até varios. Están formados por dúas brácteas, as glumelas, que pechan á flor. Nalgúns xéneros os antecios despréndense cun fragmento da raquiña, frecuentemente cuberto de pelos que forma o antopodio ou callus(Aristida, Stipa). Noutros xéneros con glumas caducas co antecio, hai un callus formado polo ápice do pedicelo (exemplo, en Heteropogon). A glumela inferior está xeralmente máis desenvolvida e denomínase lemma; é a bráctea onde se desenvolve a flor; a lemma é ovada ou lanceolada; comprimida lateral ou dorsalmente, cunha ou varias nervaduras, aguda ou obtusa, mútica ou cunha ou máis arestas apicais ou dorsais. A glumela superior ou pálea é o profilo situado entre a flor e a raquilla; xeralmente é menor que a lemma e está máis ou menos cuberta polos bordos desta; adoita ser de consistencia membranosa, e posúe polo común dúas nervaduras prominentes formando dúas carenas. A pálea pode estar atrofiada ou, mesmo, faltar.
Flor. A flor é espida (carece de cáliz e corola), mais está acompañada xeralmente por dúas (raramente tres) pequenas pezas escamosas traslúcidas, chamadas lodículas, que constitúen un resto de perianto. A flor pode ser hermafrodita ou unisexual. En moitos xéneros os antecios levan flores hermafroditas, salvo os superiores que son masculinas. En cambio, en moitas Paníceas, hai un antecio inferior masculino e un superior hermafrodita. Outras veces hai espiguillas masculinas e espiguillas femininas na mesma planta (o caso de Zea mays, especie diclino monoica) ou en plantas distintas (exemplo, no subxénero Dioicopoa de Poa, constituído por especies diclino dioicas). As lodículas semellan ser restos dun perianto trímero e aínda existen 3 lodículas nalgúns xéneros de bambúseas e nalgunhas especies de Stipa. A turxencia das lodículas determina a apertura do antecio permitindo a saída ao exterior dos estames e estigmas (momento que se chama floración). A morfoloxía das lodículas ten importancia sistemática. Así mesmo, as flores adoitan posuír algún destes dous tipos de compresión, importantes taxonómicamente: ou están comprimidas lateralmente (de forma que a lemma e a pálea obsérvense a ambos os dous lados da cara comprimida), ou están comprimidas ventralmente (de xeito que cada cara comprimida posúa á lemma ou á pálea).
Androceo. O androceo nas gramíneas é cíclico e en xeral trímero (é dicir, está composto por tres estames, ou un múltiplo de tres).Flores en antesis de Holcus mollis, obsérvense os estigmas plumosos e os estames. O número máis frecuente de estames é 3, mais en moitas especies de oríceas e de bambúseas hai de 6 a 9 (ás veces máis). En troques, en Imperata brasiliensis hai un só estame e dous no xénero Anthoxanthum. As anteras son biloculares e insérense sobre filamentos estreitos e máis ou menos longos. Os grans de pole son relativamente pequenos, con paredes moi tenues, liviáns, teñen un só poro xerminativo (dinse monoporados) e están adaptados a ser levados polo vento desde os estames até os estigmas doutras plantas (a polinización é anemófila).
Xineceo. O xineceo está formado por un ovario globoso, piriforme ou fusiforme, xeralmente bicarpelar, unilocular, con dous estilos curtos e estigmas plumosos. Nalgunhas bambúseas o ovario é tricarpelar e leva 3 estilos. Cando hai 3 carpelos, a peza adaxial é fértil (Kircher 1986). En Euchlaena e Zea existe un só estilo acrescente. Dentro do ovario, xineceo|súpero, hai un só óvulo anfítropo ou semianátropo, subapical ou case basal, de placentación parietal. Algúns pastos teñen óvulos péndulos, átropos. A parede do macrosporanxio pode ser delgada ou grosa. En moitas especies de gramíneas existen flores cleistógamas en inflorescencias reducidas situadas nas axilas das follas inferiores.[2][3]
Esquema dunha cariopse de gramínea, amosando o endosperma, o embrión e diversas estruturas embrionais: o escutelo ou escudete, a plúmula, a coleorriza, e o nó cotiledonar. Rodeando a todo o endosperma apréciase a capa de aleurona e ao outro extremo do embrión, o cepillo.Cariopses de arroz (Oryza sativa) en corte lonxitudinal, os embrións áchanse tinguidos de cor azul, o endosperma en branco.
O froito ou gran das gramíneas é unha cariopse, froito seco, cunha semente cuxa testa está ligada co pericarpo formando unha envoltura moi delgada. Esta envoltura encerra ao embrión e ao endosperma. Este froito é basicamente unha variante do aquenio, aínda que se pode atopar unha variedade de froitos na familia (ver por exemplo Werker 1997). Nalgúns xéneros como Zizianopsis ou Eleusine, o pericarpio non está soldado coa semente, de modo que o froito é un aquenio (ou un utrículo segundo outros autores). Nalgunhas bambúseas o froito é unha noz ou unha baga, mentres no xénero Sporobolus o pericarpio deixa saír á semente cando se embebe en auga. Moitos xéneros, como Aristida, Stipa, Piptochaetium,Oryza e case todas as Paníceas posúen cariopses que se desprenden da planta envolvidos pola lemma e pola pálea. Nas Andropogóneas son as glumas as que persisten pechando á cariopse. En Pennisetum e Cenchrus despréndese toda a espiguilla rodeada dun invólucro de porcas ou de espiñas. A forma da cariopse varía moito segundo os xéneros, podendo ser case circular como en Briza, oblonga como en Hordeum, lanceolada como en Poa até case lineal, como en Vulpia.
Na parte inferior da cariopse vista dorsalmente, vese o embrión máis ou menos elíptico cuberto polo pericarpio transparente. Do outro lado, correspondente ao asuco ou sutura carpelar, distínguese, tamén por transparencia, a mácula fiar ou fío (ou zona de unión da semente co carpelo), que pode ser punctiforme, como en Poa e nas Paníceas, ovada, como en Briza subaristata, ou lineal, como en Hordeum, Vulpia ou Festuca.
Embrión. O embrión das gramíneas é estruturalmente moi complicado e consta da plántula unida á súa cotiledón laminar, altamente modificado, chamado escudete. O cotiledón é delgado, parenquimatoso, levando no seu parte exterior unha capa de células epiteliais que durante a xerminación botan encimas. A planta consta dun nó cotiledonar, onde se insere o cotiledón, unha xema cuberta dun coleoptilo e unha radícula envolvida por outra coleorriza. En moitos xéneros na parte externa do nó cotiledonar hai unha escama diminuta, o epiblasto, que para algúns autores constitúe un resto dun segundo cotiledón, mentres que outros consideran que se trata dun apéndice da coleorriza.[2][3]
O tamaño e o número de cromosomas teñen grande importancia na sistemática das Gramíneas. Hai dous tipos cromosómicos extremos: o tipo festucoide caracterizado por presentar cromosomas grandes e número básico predominantemente x=7 e o tipo panicoide con cromosomas pequenos e números básicos predominantes x=9 e x=10. O tipo festucoide atópase en case todas as tribos da subfamilia Festucoideae (sen.: Pooideae), con algunhas excepcións. Por exemplo, a tribo Stipeae desta subfamilia posúe cromosomas pequenos e números básicos x=9, 10, 11, 12, 14, 16 e 17. Os restantes subfamilias das gramíneas presentan o tipo cromosómico panicoide, con cromosomas pequenos e predominio do número básico x=9 e 10. Nas Bambusóideas, ehrhartóideas e arundinóideas os cromosomas son pequenos e o número básico é x=12. Na subfamilia Danthonioideae preséntanse cromosomas de tamaño intermedio e número básico x=6 e 7. A subfamilia Chloridoideae presenta cromosomas pequenos e varios números básicos, x=7, 8, 9, 10, 11, 12 e 14. As Panicóideas sempre teñen cromosomas pequenos, con números básicos x=9 ou x=10, aínda que existen especies con outros números básicos, que varían desde x=4 a x=19.[3]
As gramíneas son morfoloxicamente distintas de calquera outra familia de plantas e, ademais, son moi diversas en canto a morfoloxía e hábito de crecemento. As diferentes especies de gramíneas - como se describiu na sección previa - difiren nos seus tamaños e números cromosómicos. Así mesmo, difiren no tamaño (ou contido de ADN) dos seus xenomas. O xenoma do arroz, por exemplo, é máis de 11 veces máis pequeno que o xenoma da cebada, a pesar de que ambas as especies son diploides e aparentan ter a mesma complexidade morfolóxica e fisiolóxica.[5] O contido de xenes das diferentes especies de gramíneas, no entanto, non varía tan amplamente como o contido de ADN total. O arroz e a cebada, novamente, non difiren máis que en 2 veces no número media de fragmentos de restrición que hibridan coas mesmas sondas.[6] A maior parte das diferenzas no tamaño do xenoma entre especies de gramíneas débese a diferenzas no ADN repetitivo. Os xenomas máis grandes, como os de cebada ou trigo, están compostos nun 75% de ADN repetitivo, mentres que os xenomas máis pequenos, como o do arroz, só conteñen menos do 50% de ADN altamente repetitivo. Máis aínda, determinouse que boa parte dese ADN repetitivo está composto de retrotransposóns insertos entre os xenes.[7][8]
A maior parte das diferenzas no tamaño do xenoma entre especies de gramíneas débese a diferenzas no ADN repetitivo. Os xenomas máis grandes, como os de cebada ou trigo, están compostos nun 75% de ADN repetitivo, mentres que os xenomas máis pequenos, como o do arroz, só conteñen menos do 50% de ADN altamente repetitivo. Máis aínda, determinouse que boa parte dese ADN repetitivo está composto de retrotransposóns insertos entre os xenes.[7][8]
Os estudos de mapeo xenómico en moitas especies de gramíneas empregando as mesmas sondas de ADN amosaron que non só o contido de xenes está moi conservado, senón tamén a orde dos xenes dentro dos cromosomas. A extensa conservación no contido de xenes e na orde dos mesmos entre o millo e o sorgo non é inesperada xa que ambas as especies "só" contan con 15 a 20 millóns de anos de evolución independente. No entanto, similares observacións para o arroz e o millo, as cales afástaronse hai de 60 a 80 millóns de anos, indican que todas as especies da familia proveñen dun mesmo antepasado común e que todas elas conservan un mesmo repertorio de xenes no mesmo orde aproximado.[9][10][11]
Os grandes arranxos xenómicos que diferencian entre si a todas as gramíneas son o resultado de investimentos ou duplicacións cromosómicas que involucran a maior parte dos brazos cromosómicos.[12]
A maioría, senón todas, as gramíneas son poliploides. Baseados no suposto que todos os xéneros e familias que presentan un número cromosómico básico x=12 son derivados de devanceiros que sufriron duplicacións cromosómicas durante a súa evolución e que as subfamilias de gramíneas máis primitivas (Anomochlooideae, Pharoideae, e Puelioideae) teñen un número cromosómico básico x=12, pódese deducir que o devanceiro das gramíneas xa era un poliploide. Séguese, ademais, que todas as gramíneas que se clasifican como diploides son, en realidade, paleopoliploides (ou sexa, poliploides antigos que presentan herdanza disómica.[13] e cuxos proxenitores non poden ser identificados mediante ferramentas citogenéticas ou marcadores moleculares).[14][15]
A familia contén máis do 60% de especies, distribuídas en todos os clados, que se clasifican como neopoliploides, ou sexa que sufriron un ciclo adicional de duplicación xenómica. Nestas especies, os xenomas duplicados non han divergido moito do xenoma dos seus devanceiros e o seu número de cromosomas e comportamento citológico durante a meiose son indicativos da duplicación cromosómica que os orixinou. A maioría destes neopoliploides (máis do 65%) derivaron de cruzamientos interespecíficos ou intergenéricos polo que se lles clasifica como alopoliploides.[14]
As hemicelulosas e os polisacáridos de pectinas da parede celular primaria dos pastos son moi diferentes dos das demais espermatófitas, tanto en estrutura como nas particularidades da composición dos xiloglucanos. Os polisacáridos son menos ramificados que en todas as demais familias de plantas, aínda que esta afirmación está baseada nunha mostraxe aínda escasa de especies. As poáceas poden ser cianoxenéticas ou non. Cando son cianoxenéticas, os compostos cianoxenéticos son derivados da tirosina. Poden presentar alcaloides (ás veces isoquinolina, pirrolizidina e indol). Raramente pode haber proantocianidinas e cianidinas, en cantidades traza, e só en representantes das subfamilias Panicoideae e Chloridoideae. Os flavonoides acháronse só nalgúns xéneros, BoutelouaGlyceria e Melica,[16] cando están presentes son quercetina, ou kaempferol xunto con quercetina.
Unha xeneralización acerca do modo de reprodución das gramíneas é que os membros desta familia son plantas hermafroditas, que presentan fertilización cruzada (son alógamos) e se polinizan polo vento. Obviamente, unha familia con preto de 10.000 especies conta con moitas excepcións a esta regra, as cales se describen a continuación.
Este tipo de sistema reprodutivo, no cal existen plantas femininas e plantas masculinas, non é moi frecuente nas gramíneas. Só 18 xéneros son dioicos ou presentan especies dioicas, sendo Poa o máis coñecido deles. De feito, as especies dioicas de Poa inclúense nun subxénero afastado, Dioicopoa.[17]
Este sistema reprodutivo describe o feito de que nas poboacións naturais dunha especie coexisten individuos femininos e individuos hermafroditas. Esta condición é bastante rara nas gramíneas. Bouteloua chondrosioides e algunhas especies do subxénero Andinae de Poa son xinodioicas, aínda que Cortadeira é o exemplo máis conspicuo.[17]
Neste sistema os sexos están separados espacialmente pero no mesmo individuo, ou sexa, cada planta presenta inflorescencias femininas e masculinas. Zea, Humbertochloa, Luziola, Ekmanochloa e Mniochloa son exemplos de xéneros con especies monoicas. Moito máis común entre as gramíneas son as especies andromonoicas, unha condición moi común nas andropogóneas e paníceas. Nas primeiras, os dous sexos preséntanse en espiguillas diferentes de pares heterógamos. Un par heterógamo de espiguillas consiste xeralmente nunha espiguilla sésil, cunha flor neutra e outra hermafrodita, e unha espiguilla pedicelada, cunha flor neutra e outra masculina. Nas espiguillas bifloras das Paniceas, en troques, a flor inferior é xeralmente masculina ou neutra, e o superior é hermafrodita. Algúns dos xéneros que exemplifican este tipo de sistema son Alloteropsis,Brachiaria,Cenchrus,Echinochloa,Melinis,Oplismenus,Panicum,Setaria,Whiteochloa, e Xyochlaena. Algunhas especies dentro destes xéneros poden ter só flores hermafroditas xa que a flor inferior é sempre neutra, raramente ambas as flores son hermafroditas. á parte das paníceas e as andropogóneas, Arundinelleae é outra tribo con especies andromonoicas. No resto da familia, as especies andromonoicas atópanse moi esporádicamente, por exemplo en Arrhenatherum,Hierochloe e Holcus.[17]
A gran maioría das especies de gramíneas son hermafroditas, no entanto, frecuentemente son incapaces de producir sementes cando o pole dunha planta poliniza os seus propios estigmas. Isto débese a que unha gran parte das especies da familia presentan autoincompatibilidade, de tipo gametofítica e debido á acción de dous xenes independentes (chamados S e Z) con varios alelos cada un. Este sistema de autoincompatibilidade foi observado en varios xéneros da familia (Festuca,Secale,Lolium,Hordeum,Dactylis, entre moitos outros) e non é perfectamente eficiente. De feito, da maioría das especies autoincompatibles pode obterse unha proporción - aínda que reducida - de sementes ao autofecundar unha planta.[17][18]
A autopolinización e a autofecundación están moi distribuídas entre as gramíneas. En xeral, é un mecanismo máis común entre as especies anuais que entre as perennes e, decididamente, moito máis frecuente entre as especies colonizadoras. Este mecanismo determinouse en aproximadamente 45 xéneros de gramíneas, entre os cales se achan xéneros economicamente moi importantes como Triticum,Oryza,Secale,Avea,Agropyron e Lolium. Unha condición de autogamia extrema é a cleistogamia. Este último sistema está distribuído en máis de 70 xéneros pertencentes a 20 tribos de gramíneas.[17][19]
A apomixia defínese como a reprodución asexual a través de sementes. Neste sistema reprodutivo os embrións desenvólvense por mitose a partir dunha oosfera non reducida sen que teña lugar a fecundación. Noutras palabras, cada embrión producido é xeneticamente idéntico a plánta nai. Nas gramíneas, a apomixia foi descrita por primeira vez en 1933 nunha especie de Poa. Desde aquel momento identificouse este mecanismo en centos de especies de poáceas, particularmente nas Paníceas e nas Andropogóneas. Algúns dos xéneros que presentan especies apomícticas son Apluda,Capillipedium,Heteropogon,Themeda,Sorghum, Bothriochloa,Dichanthium, Cenchrus,Setaria e Paspalum.[17][20][21]
↑Judd, W. S., Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. (2002), «Poaceae» Plant systematics: a phylogenetic approach, Sinauer Axxoc, 287-292. ISBN 0-87893-403-0.
↑ 2,02,12,22,3Parodi, L.R. 1987. Gramíneas. En: Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires, PP: 108-182
↑ 3,03,13,23,33,4Cabrera, A. L., Cámara Hernández, J., Caro, J.; Covas, G.; Fabris, H., Hunziker, J., Nicora, E., Rugolo, Z., Sánchez, E., Torres, M. (1970), «Gramineae, parte general.» Flora de la Provincia de Buenos Aires: Gramíneas., Colección Científica del INTA. Tomo IV, parte II., 1-18.
↑Carenada é un termo que se emprega en botánica para referirse a calquera órgano da planta que presenta a forma dunha quilla ou carena. Ref.: Parodi, L.R. 1987. Gramíneas. En: Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Segundo Volumen. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires, PP: 1114.
↑Kurata, N., G. Moore, Y. Nagamura, T. Foote, M. Yano, Y. Minobe, and M.D. Gale. 1994. Conservation of genome structure between rice and wheat. BioTechnology 12: 276-278 resumo en inglés
↑ 7,07,1Flavell, R.B., M.D. Bennett, J.B. Smith, and D.B. Smith. 1974. Genome size and proportion of repeated nucleotide sequence DNA in plants. Biochem. Genet. 12: 257-269
↑ 8,08,1SanMiguel, P., A. Tikhonov, Y.-K. Jin, N. Motchoulskaia, D. Zakharov, A. Melake-Berhan, P.S. Springer, K.J. Edwards, Z. Avramova, and J.L. Bennetzen. 1996. Nested retrotransposons in the intergenic regions of the maize genome. Science 274: 765-768 resumo en inglés
↑Clark, L.G., W. Zhang, and J.F. Wendel. 1995. A phylogeny of the grass family (Poaceae) based on ndhF sequence data. Syst. Bot. 20: 436-460
↑Bennetzen, J.L. and M. Freeling. 1993. Grasses as a single genetic system: Genome composition, collinearity and compatibility. Trends Genet. 9: 259-261
↑Bennetzen, J.L. & Freeling, M. 1997.
The Unified Grass Genome: Synergy in Synteny
Genome Res.7: 301 - 306.Artigo en inglés
↑Moore, G., T. Foote, T. Helentjaris, K. Devos, N. Kurata, & M. Gale. 1995. Was there a single ancestral grass chromosome? Trends Genet.
↑Herdanza disómica é o tipo de herdanza que presentan os organismos diploides, os que levan só dous xogos de cromosomas. Aplicado a un poliploide indica que, desde o punto de vista xenético e pese a estar constituido por varios xogos de cromosomas, ese organismo compórtase como un diploide.
↑ 14,014,1Levy, A. & Moshe Feldman. 2002. The Impact of Polyploidy on Grass Genome Evolution Plant Physiol. 130: 1587-1593 Artigo en inglés
↑A paleopoliploidía pódese detectar só a través de ferramentas bioinformáticas bastante sofisticadas que revelan a similitude entre xenes que se afastaron millónes de anos atrás.
↑Baumann, U., Juttner, J., Bian, X. & P. Langridge. 2000. Self-incompatibility in the Grasses. Annals of Botany 85 (Supplement A): 203-209. Artigo en inglés
↑Campbell, C., J. Quinn, G. Cheplick, & T J Bell. 1983. Cleistogamy in Grasses. Annual Review of Ecology and Systematics Vol. 14: 411-441.