Segmentación (zoología)
Se denominan segmentación o clivaje al proceso embriológico temprano que consiste en una serie de divisiones celulares (Mitosis) del óvulo fecundado (cigoto) que se producen antes de la gastrulación y que se relacionan con la morfología del huevo y en particular con la cantidad de vitelo que contiene. Las células resultantes de la división del cigoto se denominan blastómeros y forman una masa compacta llamada mórula; a partir de ésta se forma la blástula y posteriormente la gástrula. El clivaje resulta de dos procesos: cariocinesis (división del núcleo cuyo agente mecánico es el huso mitótico) y citocinesis (división celular cuyo agente mecánico es el anillo contráctil).
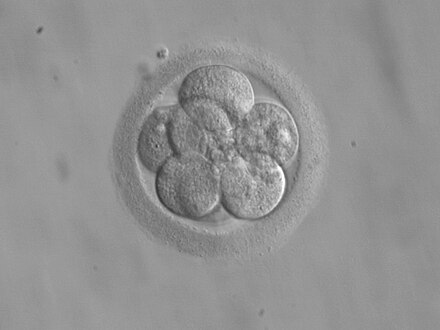
Tipos de segmentación
editarEl tipo son 19 de segmentación está estrechamente relacionado con la morfología del cigoto, y en particular con la cantidad y la disposición del vitelo, lo cual, a su vez, depende de las características del óvulo (el espermatozoide no contiene substancias de reserva). La condición del huevo según la cantidad y distribución de la proteína vitelina dentro del citoplasma puede clasificarse según cantidad en:
- Oligolecito: Baja cantidad de vitelo (erizos, gusanos, moluscos y mamíferos).
- Mesolecito: Mediana cantidad de vitelo (anfibios).
- Polilecitos: Gran cantidad de vitelo (aves, insectos, reptiles y peces).
Otra clasificación es según distribución:
- Isolecito: Vitelo distribuido uniformemente (erizos y mamíferos).
- Telolecito: Vitelo desplazado al polo vegetal (peces, anfibios, aves).
- Centrolecito: Vitelo acumulado en el centro (insectos).
Se distinguen dos tipos básicos de segmentación, aunque hay que tener en cuenta que entre ellos hay otros muchos intermedios, según la cantidad de vitelo del huevo y su disposición. En otros casos, como en el huevo alecítico de los mamíferos placentarios, al perder secundariamente el vitelo, inicia su segmentación de modo holoblástico y la continua de manera meroblástica.
Segmentación total u holoblástica
editarEs típica de los huevos que tienen poco vitelo y en cada división participa todo el cigoto. Por tanto, se da básicamente en huevos isolecíticos y en los heterolecíticos. A su vez, la segmentación total puede ser igual o desigual.
- Segmentación total igual. Es propia de algunos huevos isolecíticos; todos los blastómeros tienen las mismas dimensiones.
- Segmentación total desigual. Es el tipo de segmentación total más extendido, ya que se da en la mayoría de huevos isolecíticos y en todos los heterolecíticos. En ella se distinguen dos tipos de blastómeros, los micrómeros, pequeños y situados en el polo animal, y los macrómeros, más grandes y situados en el polo vegetativo.
Segmentación parcial o meroblástica
editarSe da cuando el huevo contiene mucho vitelo y solo se divide el polo animal, formándose un pequeño casquete de células sobre el vitelo. La segmentación no afecta al polo vegetativo y la mayor parte del vitelo queda sin segmentar. Este tipo de segmentación presenta dos modalidades:
- Segmentación parcial discoidal. Es característica de los huevos telolecíticos. La división solo afecta a un disco citoplasmático próximo al polo animal y la parte inferior (polo vegetativo) no se segmenta; a continuación se forma un disco de blastómeros, el blastodermo o blastodisco, a partir del cual se formará el embrión, que reposará sobre la masa vitelina. Se da en peces, reptiles y aves, entre otros.
- Segmentación parcial superficial. Es típica de los huevos centrolecíticos. El núcleo se divide repetidas veces sin que aparezcan límites celulares definidos dentro de la masa vitelina; luego, los núcleos emigran al citoplasma periférico y se disponen formando una capa sincitial; finalmente surgen los límites celulares que delimitan un blastodermo periférico alrededor del vitelo central no segmentado. Ocurre típicamente en artrópodos.
Otra clasificación de la segmentación se fundamenta en el destino de los blastómeros:
Segmentación indeterminada
editarSe da principalmente en los deuterostomados. Cada célula producida por la segmentación tiene el potencial de desarrollarse en un embrión completo. Debido a ello es posible es desarrollo de técnicas de laboratorio como ser la clonación de individuos.
Segmentación determinada
editarSe da, sobre todo, en protostomados. Cada blastómero, separado experimentalmente, solamente origina una parte determinada del animal, la misma que habría dado en el caso de no haberse separado.
Otra diferencia en el tipo de segmentación consiste en el plano según el cual se dividen los blastómeros y, por tanto, en la disposición que éstos adoptan. Así, se distinguen dos tipos principales, que solo se ponen claramente de manifiesto en la segmentación holoblástica:
Segmentación radial
editarLa segmentación radial es indeterminada y se da principalmente en deuterostomados (por ejemplo, los equinodermos, hemicordados y cordados). Durante la segmentación se establece una alternancia regular de surcos de división meridianos y longitudinales; los primeros delimitan regiones de blastómeros verticales, mientras que los segundos conducen a la formación de blastómeros dispuestos en filas horizontales. Así se establece una simetría radial respecto del eje de polaridad de la blástula.
Segmentación espiral
editarLa segmentación espiral es determinada y muy característica de los protostomados del clado Spiralia (por ejemplo, los platelmintos, la mayoría de los moluscos, anélidos, nemertinos, etc.). Los planos de segmentación son oblicuos en relación al eje del huevo. Los blastómeros hijos se colocan entre los blastómeros padres y quedan dispuestos en capas horizontales, con los blastómeros en posición alternada, por lo que se disponen en espiral.
Estos dos tipos de segmentación representan una dicotomía ancestral, la divergencia evolutiva de los metazoos bilaterales en dos linajes separados, protóstomos y deuteróstomos.
Segmentación en diferentes grupos de organismos
editarErizo de mar
editarPresentan clivaje holoblástico radial. La primera y segunda segmentaciones son meridionales (perpendiculares), es decir que los surcos de segmentación pasan a través de los polos vegetal y animal. La tercera segmentación es ecuatorial (perpendicular a las dos primeras) y separa entre sí los hemisferios animal y vegetal. Durante la cuarta segmentación, las cuatro células del nivel animal se dividen meridionalmente en ocho blastómeras con el mismo volumen (mesómeras), y el nivel vegetal experimenta segmentación ecuatorial desigual y produce cuatro células grandes (macrómeras) y cuatro pequeñas (micrómeras). Durante la quinta segmentación, las ocho mesómeras animales se dividen ecuatorialmente y producen dos niveles: an1 y an2, uno sobre el otro. Las macrómeras se dividen meridionalmente y forman un nivel de ocho células debajo de an2. Las micrómeras se dividen más tarde y producen un pequeño grupo debajo del nivel más grande. En la sexta segmentación, las células del hemisferio animal se dividen meridionalmente y las vegetales ecuatorialmente. En la séptima segmentación, las células animales se dividen ecuatorialmente y las vegetales meridionalmente. En ese momento, el embrión es una blástula de 128 células y las divisiones se vuelven menos frecuentes (Gilbert, 2005).
Caracol
editarPresentan clivaje holoblástico espiral. Su plano de segmentación es oblicuo formando una disposición en espiral de las blastómeras. Las primeras dos divisiones son casi meridionales y producen cuatro grandes macrómeras. En las siguientes divisiones desde cada macrómera surge una micrómera en su polo animal. Cada cuarteto de micrómeras es desplazado a la derecha o izquierda de su macrómera hermana creando el patrón espiral. Durante la tercera división, las macrómeras forman, cada una, dos células hijas (otra macrómera y una micrómera). En la cuarta división, desde la macrómera se producen una macrómera y una micrómera, mientras que de la micrómera se producen dos micrómeras más, formando el segundo cuarteto de micrómeras. El primer cuarteto formará estructura de la cabeza y el segundo cuarteto formará el estatocisto (órgano del equilibrio) y la concha (Gilbert, 2005).
Tunicados
editarPresentan clivaje holoblástico bilateral. El primer plano de clivaje establece el eje más temprano de simetría en el embrión separándolo en sus futuros lados derecho e izquierdo. Cada división sucesiva se orienta en relación con este plano de simetría y la mitad del embrión formada sobre uno de los lados del primer plano de división es la imagen de espejo de la mitad del embrión sobre el otro lado. La segunda segmentación también es meridional, pero no pasa a través del centro del huevo, sino que se generan dos células anteriores grandes y dos células posteriores pequeñas. En el estadio de 64 células se forma un pequeño blastocele y comienza la gastrulación (Gilbert, 2005).
Caenorhabditis elegans
editarPresentan clivaje holoblástico rotacional. Cada división asimétrica produce una célula fundadora (AB, MS, E, C, D), que produce descendientes diferenciados y una célula madre (P1-P4). En la primera división, el surco de segmentación se encuentra dispuesto asimétricamente a lo largo del eje anteroposterior y más cerca de los que será el polo posterior. Esa división forma una célula fundadora anterior y una célula madre posterior. En la segunda división, esa célula fundadora se divide ecuatorialmente, mientras que la célula madre se divide meridionalmente para producir una célula fundadora anterior y una célula madre posterior. El linaje de células madre siempre se dividirá meridionalmente para crear una célula fundadora anterior y una célula madre posterior que continuará con el linaje de células madre (Gilbert, 2005).
Mamíferos
editarPresentan clivaje holoblástico rotacional, pues la primera segmentación es una división meridional y durante la segunda segmentación, una de las dos blastómeras se divide meridionalmente mientras que la otra se divide ecuatorialmente. La división celular temprana es asincrónica, es decir que las blastómeras no se dividen al mismo tiempo. Además, las blastómeras durante el estadio de ocho células forman una organización laxa con abundante espacio entre sí, llamada compactación (Gilbert, 2005).
Anfibios
editarPresentan clivaje radialmente asimétrico y holoblástico. Tienen mucho vitelo en el polo vegetal que causa el clivaje desplazado. La primera división es meridional y comienza en el polo animal y se extiende al polo vegetal, la segunda división también es meridional y perpendicular a la primera. La tercera división es ecuatorial desplazada hacia el polo animal. El surco de segmentación divide al embrión en cuatro blastómeras animales pequeñas (micrómeras) y cuatro blastómeras vegetales grandes (macrómeras). El embrión es considerado mórula durante el estadio de 16-64 células y es considerado blástula en el estadio de 128 células, cuando comienza a observarse el blastocele (Gilbert, 2005).
Pez cebra
editarPresentan clivaje meroblástico discoidal, debido a que la segmentación sucede solo en el blastodisco (región delgada del citoplasma libre de vitelo en el polo animal) y solo esta región llegará a ser el embrión. El cigoto tiene una estructura con forma de pera con un blastodisco apical, pues el citoplasma no vitelínico está restringido al polo animal. Las primeras doce divisiones son sincrónicas y rápidas con segmentaciones meridionales y ecuatoriales, formando un montículo de células sobre el polo animal (blastodermo). En el momento de transición hacia blástula media (cerca de la décima división celular), se pueden distinguir tres poblaciones celulares diferentes: la primera es la capa sincicial vitelínica que se forma cuando las células del borde vegetal del blastodermo se fusionan con la célula vitelínica subyacente produciendo un anillo de núcleos dentro de la parte citoplasmática de la célula vitelínica, y será importante para dirigir algunos de los movimientos de gastrulación; la segunda es la capa de la envoltura que es formada por las células más superficiales del blastodermo , y se convertirá finalmente en el peridermo (cubierta protectora extraembrionaria liberada durante el desarrollo tardío); la tercera son las células profundas encontradas entre la capa de la envoltura y la capa sincicial vitelínica, y darán origen al embrión propiamente dicho (Gilbert, 2005).
Pollo
editarPresentan clivaje meroblástico discoidal. El primer surco de segmentación aparece centralmente en el blastodisco y otros surcos de segmentación siguen para crear un blastodermo de una sola capa. Las segmentaciones ecuatorial y vertical dividen al blastodermo en un tejido de cinco a seis capas celulares de espesor. Entre el blastodermo y el vitelo hay un espacio denominado cavidad subgerminal, el cual es creado cuando las células del blastodermo absorben agua desde la albúmina (clara del huevo) y secretan el fluido entre ellas y el vitelo.
Drosophila
editarPresenta un tipo de clivaje superficial, pues tienen una gran masa de vitelo en la parte central que restringe el clivaje al borde o aro citoplasmático del cigoto. El núcleo del cigoto experimenta muchas divisiones mitóticas, se producen 256 núcleos mediante ocho divisiones nucleares. Los núcleos migran a la periferia del huevo y se siguen dividiendo. Durante la novena división, cerca de cinco núcleos alcanzan la superficie del polo posterior del embrión y llegan a ser incluidos por las membranas celulares formando las células polares que originan los gametos en el adulto. La mayoría del resto de núcleos alcanzan la periferia en el décimo ciclo y después presentan cuatro divisiones más (el embrión se denomina sincicial, pues los núcleos están contenidos dentro del mismo citoplasma). Después de la división número trece, la membrana plasmática del huevo se pliega hacia dentro entre los núcleos separando cada núcleo somático en célula individual, creando así el blastodermo celular en el que todas las células están organizadas en una capa única rodeando el centro del huevo que contiene el vitelo. El blastodermo celular consiste de 6000 células y se produce luego de una serie de invaginaciones por parte de las membranas celulares (Gilbert, 2005).
Xenopus laevis
editarVéase también
editarBibliografía
editar- Altaba, C. R. et al., 1991. Invertebrats no artròpodes. Història Natural dels Països Catalans, 8. Enciclopèdia Catalana, S. A., Barcelona, 598 pp. ISBN 84-7739-177-7
- Gilbert, S. 2005. Biología del Desarrollo, 7a Edición. Sinauer Assoc, Sunderland.
- Hickman, C. P., Ober, W. C. & Garrison, C. W., 2006. Principios integrales de zoología, 13ª edición. McGraw-Hill-Interamericana, Madrid (etc.), XVIII+1022 pp. ISBN 84-481-4528-3
- Kozlowski, R. N., Tyler M. S. & Bagley, C. 2008. Stages in embryonic development of Drosophila melanogaster. Developmental biology. BioMediaLab. Rescatado de http://www.devbio.net/node/44.
- Schierenberg, E. Development and Evolution. Development and evolution in nematodes. Rescatado de https://web.archive.org/web/20110102165532/http://www.uni-koeln.de/math-nat-fak/zoologie/expmorph/agschier/schier2e.html.
- Urven, L. E., Yabe, T. & Pelegri, F. 2006. A role for non-muscle myosin II function in furrow maturation in the early zebrafish embryo. Journal of Cell Science, Vol. 119, 4342-4352.
- Wilt, F. H. & Hake, S. C. 2004. Principles of Developmental Biology. W.W. Norton & Company, Inc.
Datos: Q376172
Multimedia: Cleavage (embryo) / Q376172
- ↑ Gilbert, Scott F. (2006). Biología del Desarrollo, 7ª. edición, Buenos Aires: Panamericana. ISBN 950-06 0869-3