Apatosaurus
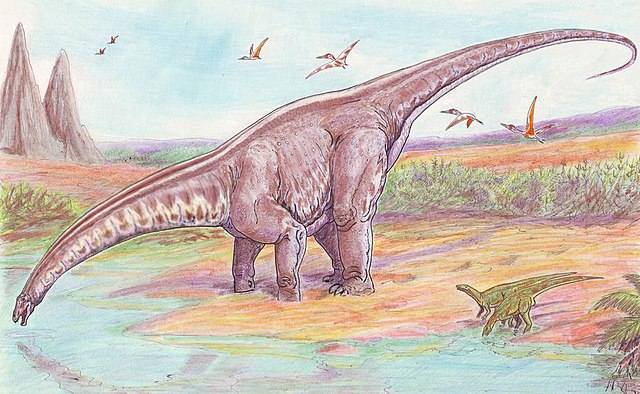
Apatosaurus (gr. "lagarto engañoso") es un género con dos especies conocidas extintas de dinosaurio saurópodo diplodócido, que vivió a finales del período Jurásico, hace aproximadamente entre 155,7 y 150,8 millones de años, en el Kimmeridgiense, en lo que hoy es Norteamérica.[1] Apatosaurus, fue el primer gran saurópodo cuyo esqueleto completo se expuso al público. Othniel Charles Marsh describió y nombró la primera especie conocida, Apatosaurus ajax en 1877 y una segunda especie, Apatosaurus louisae, fue descubierta y nombrada por William H. Holland en 1916. Vivieron hace unos 152 a 151 millones de años, durante el Titoniano temprano, y se conocen a partir de fósiles en la formación Morrison de la actual Colorado, Oklahoma y Utah, en Estados Unidos. Apatosaurus tenía una longitud entre 21 a 22,8 metros, y una masa entre 16,4 a 22,4 toneladas. Unos ejemplares indican una longitud máxima de 11-30 % mayor que la media y una masa entre 32,7 a 72,6 toneladas.
| Apatosaurus | ||
|---|---|---|
| Rango temporal: 155,7 Ma - 150,8 Ma Jurásico Superior | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| Superfamilia: | Diplodocoidea | |
| Familia: | Diplodocidae | |
| Subfamilia: | Apatosaurinae | |
| Género: |
Apatosaurus Marsh, 1877 | |
| Especie tipo | ||
|
Apatosaurus ajax Marsh, 1877 | ||
| otras Especies | ||
| ||
Las vértebras cervicales de Apatosaurus son menos alargadas y más pesadamente construidas que las de Diplodocus, un diplodócido como Apatosaurus, y los huesos de la pierna son mucho más robustos a pesar de ser más largos, lo que implica que Apatosaurus era un animal más robusto. La cola se mantenía por encima de la tierra durante la locomoción normal. Apatosaurus tenía una sola garra en cada extremidad anterior y tres en cada uno de los miembros posteriores. El cráneo de Apatosaurus, que se creía que era similar al de Camarasaurus, es mucho más similar al de Diplodocus. Apatosaurus era un herbívoro generalista que probablemente llevaba su cabeza elevada. Para aligerar sus vértebras, Apatosaurus tenía sacos de aire que hicieron los huesos internos llenos de agujeros. Al igual que la de otros diplodócidos, la cola puede haber sido utilizado como un látigo para crear ruidos fuertes.
Hasta 1909, el cráneo del apatosaurio se confundió con el de Camarasaurus y Brachiosaurus, cuando se encontró el holotipo de A. louisae y un cráneo completo a solo unos pocos metros de distancia de la parte frontal del cuello. Henry Fairfield Osborn no estuvo de acuerdo con esta asociación, y montó un esqueleto de Apatosaurus con un cráneo de Camarasaurus. Hasta 1970, los esqueletos de Apatosaurus fueron montados con los moldes del cráneo especulativa de Camarasaurus, cuando McIntosh mostró que los cráneos más robustos asignados a Diplodocus tenían más probabilidades de ser de Apatosaurus.
El término Brontosaurus ha sido considerado como un sinónimo más moderno de Apatosaurus y su única especie, se reclasificó como A. excelsus en 1903. Sin embargo, un estudio de 2015 concluyó que Brontosaurus era un género de saurópodo válido distinto de Apatosaurus. Sin embargo, no todos los paleontólogos están de acuerdo con esta división.
A finales del Jurásico, Apatosaurus habría vivido junto a dinosaurios como Allosaurus, Camarasaurus, Diplodocus y Stegosaurus.
Descripción
editarApatosaurus era un gran animal cuadrúpedo de cuello largo, con una larga cola en forma de látigo. Sus patas delanteras eran ligeramente más cortas que las traseras. Los apatosaurios o brontosaurios eran unos de los animales más grandes que han existido en la Tierra, alcanzaban cerca de 23 metros de largo, 4,5 de alto hasta las caderas y tenían un peso de 22 toneladas. Las vértebras cervicales eran menos alargadas y estaban más fuertemente construidas que la de Diplodocus, y los huesos de la pierna eran mucho más gruesos a pesar de ser más largos, lo que implica un animal más robusto. La cola se mantenía despegada del suelo durante la locomoción normal. Como la mayoría de los saurópodos, Apatosaurus tenía solo una garra grande en cada pata delantera, con los tres primeros dedos de la extremidad posterior con garras.[2] Para ayudar en el proceso del alimento, el Apatosaurus pudo haber tragado piedras, gastrolitos, de la misma manera que lo hacen muchos pájaros, ya que sus quijadas por sí solas no eran suficientes para masticar las resistentes fibras de las plantas.
Tamaño
editarLa mayoría de las estimaciones del tamaño de Apatosaurus se basan en el espécimen tipo de A. louisae, CM 3018 En 1936, este ejemplar se midió en 21,8 m , mediante la medición de la columna vertebral.[3] Las estimaciones varían muy poco del anterior resultado, entre unos 21 y 22,8 metros de largo y con un peso de entre 16,1 a 22,4 toneladas de peso.[4][5] Un estudio de 2015 calculó la masa corporal de algunos saurópodos sobre la base de modelos volumétricos, estimando que CM 3018 pesaría entre 21,8 y 38,2 toneladas, similar en masa a un titanosario argentino llamado Dreadnoughtus.[6] Algunos especímenes gigantescos de Apatosaurus hallados en Oklahoma representan individuos de aprox. 11–30% más largos, como por ejemplo el espécimen OMNH 1670, consistente en una vértebra dorsal de 135 cm de alto, que es exactamente un 27% más grande que el correspondiente a CM 3018, lo que sugiere una masa el doble de grande, posiblemente de entre 33 y 73 toneladas. Esto sugiere que Apatosaurus potencialmente rivalice en tamaño con los mayores titanosaurios.[7][8][9]
Vértebras
editarAl igual que las de otros saurópodos, las vértebras del cuello están profundamente bifurcadas, separando espinas neurales con un gran canal en el medio, lo que resulta en un cuello ancho y profundo.[10] La fórmula vertebral para el holotipo de A. louisae es de 15 vértebras cervicales, 10 dorsales, 5, sacras y 82 caudales. El número vértebra caudales puede variar, incluso dentro de las especies.[3] Las vértebras cervicales de Apatosaurus y Brontosaurus son más gruesas y más robustas que las de otros diplodócidos, resultaron ser más similar a las de Camarasaurus de Charles Whitney Gilmore.[3][11] Además, apoyan costillas cervicales que se extienden más lejos hacia el suelo que en los Diplodocinae, y tienen las vértebras y las costillas estrechas hacia la parte superior del cuello, haciendo la sección transversal del cuello casi triangular.[11] En Apatosaurus louisae, el complejo atlas-axis, las primeras vértebras cervicales, está casi fundida. Las costillas dorsales no están fundidas o fuertemente unidas a las vértebras, estando débilmente articulado.[3] Apatosaurio tiene diez costillas dorsales a cada lado del cuerpo.[12] Apatosaurus también tenía las costillas muy largas en comparación con la mayoría de los otros diplodócidos, dándole un pecho inusualmente profundo.[13] El gran cuello se llenó con un amplio sistema de sacos aéreos para el ahorro de peso . Apatosaurus, al igual que su pariente cercano Supersaurus, tiene espinas neurales altas, que representan más de la mitad de la altura de sus vértebras. La forma de la cola es inusual para un diplodócido; Es relativamente delgado debido a la rápida disminución de altura de las espinas vertebrales al alejarse de las caderas. Al igual que en otros diplodócidos, la cola se transformaba en una estructura en forma de látigo hacia su extremo.[3]
Miembros
editarLos huesos de las extremidades también son muy robustos.[13] Dentro de los Apatosaurinae, la escápula de A. louisae es intermedia en morfología entre las de A. ajax y Brontosaurus excelsus. Los huesos de los brazos son fuertes, por lo que el húmero de Apatosaurus se asemeja al de Camarasaurus y Brontosaurus. Sin embargo, los húmeros de Brontosaurus y A. ajax son más similares entre sí de lo que son a A. louisae. En 1936, Charles Gilmore señaló que las reconstrucciones anteriores de las extremidades anteriores de Apatosaurus proponen erróneamente que el radio y el cúbito se cruzaban, ya que en vida que estarían paralelos.[3] Apatosaurus tenía una sola garra grande en cada extremidad anterior, una característica compartida por todos los saurópodos más derivados que Shunosaurus.[3][14] Los primeros tres dedos de los pies tenían garras en cada extremidad posterior. La fórmula de falange es 2-1-1-1-1, es decir, el dedo más interior en la extremidad anterior tiene dos falanges y el siguiente tiene uno.[15] El único hueso de la garra manual es ligeramente curvo y recto truncado en el extremo anterior. La cintura pélvica incluye un robusto ilion, y el pubis e isquion fusionados. Los fémures de Apatosaurus son muy robustos, algunos de los más robustos fémures de cualquier miembro de Sauropoda. Los huesos de la tibia y peroné son diferentes de los huesos delgados de Diplodocus, pero son casi indistinguibles de los de Camarasaurus. El peroné es más largo y más delgado que la tibia. El pie de Apatosaurus tiene tres garras en los dedos más internos, la fórmula digital es 3-4-5-3-2. El primer metatarsiano es el más robusto, una característica compartida entre los diplodócidos.[3][16]
Cráneo
editarEl primer cráneo fue identificado en 1975, varias décadas después de que se le diera un nombre. El cráneo es pequeño en comparación con el tamaño del animal. Las mandíbulas estaban llenas de dientes espatulados, con forma de cincel, adaptados para una dieta herbívora.[17][10] El hocico de Apatosaurus y diplodocídos similares era redondeado, y solo Nigersaurus tenía un cráneo más cuadrado.[18] La caja craneana de Apatosaurus está muy bien conservada en espécimen BYU 17096, que también conserva gran parte del esqueleto. Un análisis filogenético encontró que la caja craneana tenía una morfología similar a los de otros diplodocídos.[19] Algunos cráneos de Apatosaurus se han encontrado todavía en articulación con sus dientes. Esos dientes que tienen el esmalte de superficie expuesta que no muestra ningún arañazos, pero presentan una textura granulada y con poco desgaste.[18]
No fue hasta 1909 que un cráneo de Apatosaurus fue encontrado durante la primera expedición, dirigida por Earl Douglass, en lo que se convertiría en el Mina Carnegie en Monumento Nacional Dinosaurio. El cráneo fue encontrado a poca distancia de un esqueleto, el espécimen CM 3018 identificado como la nueva especie A. louisae por el nombre de Louise Carnegie, esposa de Andrew Carnegie quién financió la investigación de campo para encontrar esqueletos completos de dinosaurios en el oeste de Estados Unidos. El cráneo fue designada CM 11162; era muy similar a la del cráneo de Diplodocus.[20] Otra esqueleto más pequeño de A. louisae fue encontrado cerca de CM 11162, CM 3018.[21] El cráneo fue aceptado como perteneciente a los especímenes de Apatosaurus por Douglass y el director del Museo Carnegie William H. Holland, aunque otros científicos, principalmente Osborn rechazaron esta identificación. Holland defendió su punto de vista en 1914 en un discurso ante la Sociedad Paleontológica de América, sin embargo, dejó al ejemplar del Museo Carnegie sin cabeza de montaje. Mientras que algunos pensaron que Holland estaba tratando de evitar conflictos con Osborn, otros sospechaban que Holland estaba esperando hasta que se encontraron un cráneo articulado y el cuello para confirmar la asociación del cráneo y el esqueleto.[22] Tras la muerte de Holland en 1934, el personal del museo coloca un molde del cráneo de un Camarasaurus en el montaje.[20] Mientras que la mayoría de los museos estaban usando cráneos de Camarasaurus en yeso o esculpidos en montajes de Apatosaurus, el Museo Peabody de Yale decidió esculpir un cráneo sobre la base de la mandíbula inferior de un Camarasaurus, con el cráneo basado en la ilustración del cráneo de Marsh de 1891. El cráneo también incluye fosas nasales que apuntan hacia adelante, algo diferente a cualquier dinosaurio y fenestraciones que difiere del dibujo y otros cráneos.[22]
No se reportó un cráneo de Apatosaurus en la literatura hasta la década de 1970 cuando John Stanton McIntosh y David Berman redescribieron los cráneos de los Diplodocus y Apatosaurus. Encontraron que aunque nunca publicó su opinión, Holland estaba casi con toda seguridad en lo correcto, que Apatosaurus tenía un cráneo como Diplodocus. Según ellos, los cráneos que por mucho tiempo se pensó que pertenecían a Diplodocus podría, en cambio, pertenecer a Apatosaurus. Ellos reasignaron varios cráneos a Apatosaurus basados en vértebras asociadas o estrechamente asociadas. A pesar de que el apoyo a Holland, se observó que Apatosaurus podría haber poseído un cráneo como Camarasaurus, basado en un diente desarticulado de Camarasaurus encontrado en el lugar preciso donde años antes se encontró un espécimen de Apatosaurus.[17] El 20 de octubre de 1979, después de las publicaciones de McIntosh y Berman, se montó el primer cráneo verdadero de Apatosaurus en un esqueleto en un museo, el Museo Carnegie.[20] En 1998, el cráneo de la cantera Felch que Marsh incluyó en su restauración del esqueleto de 1896 se sugirió que pertenece a Brachiosaurus.[23] En 2011, se describió al primer espécimen de Apatosaurus encontrado con el cráneo articulado con sus vértebras cervicales. Este espécimen, CMC VP 7180, se encontró que difiere tanto en el cráneo y como en el cuello de A. louisae, pero comparte muchas características de las vértebras cervicales con A. ajax.[24][25] Otro cráneo bien conservado es el espécimen BYU 17096, un cráneo bien conservado y el esqueleto, con una caja craneana bien conservada. El espécimen fue encontrado en la mina Cactus Park, al oeste de Colorado.[19]
Descubrimiento e investigación
editarEl nombre Apatosaurus fue acuñado en 1877 por Othniel Charles Marsh, profesor de Paleontología de la Universidad de Yale , sobre la base de un esqueleto casi completo, el holotipo espécimen YPM 1860, descubierto en las estribaciones orientales de las montañas rocosas en el condado de Gunnison, Colorado. La especie fue A ajax.[12][26][27] Apatosaurus significa 'lagarto engañoso', habiéndole sido dado porque los huesos del cheurón eran similares a los de un lagarto marino prehistórico, Mosasaurus. Apatosaurus proviene del griego ἀπατέλος o ἀπατέλιος (apatao) 'engañoso' y σαῦρος (saurus) 'lagarto'. El nombre de la especie "ajax" Αἴας debido al héroe griego Áyax el grande de la Guerra de Troya.[26][28] Los fósiles de este animal se han encontrado en Nine Mile Quarry y Bone Cabin Quarry, en Wyoming, y en varias localidades de Colorado, Oklahoma y Utah, en EE. UU.. Durante la excavación y transporte, los huesos del esqueleto holotipo se mezclaron con los de otro individuo descrito originalmente como Atlantosaurus immanis, como consecuencia, algunos elementos no se pueden atribuir a cualquiera de las muestra con certeza.[27] Marsh distinguió el nuevo género Apatosaurus de Atlantosaurus sobre la base del número de vértebras sacras, con Apatosaurus teniendo tres y Atlantosaurus cuatro. En 1879 describió otro hallazgo de un ejemplar mucho más completo y más grande como Brontosaurus. Marsh decidió dar este espécimen un nuevo nombre, porque las convenciones y escaso registro fósil en ese momento, las características que utilizó para distinguir los géneros y especies ahora se han encontrado ser más generalizada entre los saurópodos.[12][27] Marsh nombró la nueva especie como B. excelsus, que significa 'lagarto del trueno', del griego brontē/βροντη traducido como trueno y sauros/σαυρος como lagarto, y del latín excelsus, 'excedido en gran número', basándose en que tenía el mayor número de vértebras sacras que cualquier otro género de saurópodo conocido hasta entonces.[29] Pero durante 1903 fue descubierto que el Apatosaurus era de hecho un Brontosaurus joven, y como el nombre de Apatosaurus fue publicado primero, se juzgó que éste tenía prioridad como nombre oficial; entonces el término Brontosaurus fue relegado al ser un sinónimo más moderno.[12] El nombre Brontosaurus no fue formalmente eliminado de los expedientes de la paleontología hasta 1974.
Fue el dinosaurio más grande y completo conocido en ese entonces, careciendo solamente de la cabeza, pies, y las porciones de la cola, y fueron preparados para ser la primera exposición en público del montaje completo de un esqueleto de saurópodo, en el Museo Peabody de Historia Natural de Yale en 1905. Los huesos que faltaban fueron creados usando huesos conocidos de familiares cercanos de Brontosaurus. Los pies fueron agregados de saurópodo que se descubrieron en la misma mina, también una cola formada para asemejarse a la que Marsh creía, además de un modelo compuesto de lo que él pensaba que sería el cráneo de esta criatura masiva. Este no era el de un delicado Diplodocus,[17] sino el compuesto «con huesos más grandes, más gruesos, más fuertes en el cráneo, mandíbulas inferiores y las coronas del diente a partir de material proveniente de tres diversos lugares»,[3][12][17][30] primariamente del Camarasaurus, el único saurópodo del cual se conocía buen material craneal en aquel entonces. Este método de reconstruir los esqueletos incompletos basados en los restos más completos de dinosaurios relacionados continúa en montajes de museo y restauraciones hasta la actualidad. El montaje fue supervisado por Adam Hermann, quien no logró encontrar cráneos de Apatosaurus. Hermann se vio obligado a esculpir un cráneo. Osborn dijo en una publicación que el cráneo era "muy hipotética y basada en la de Morosaurus (ahora Camarasaurus)".[22]
A pesar de la mucha publicidad dada al esqueleto montado, que cimentó el nombre Brontosaurus en la conciencia del público, en 1903, Elmer Riggs publicó un estudio que describe un esqueleto bien conservado de un diplodócido Gran Valle cerca de Fruita , el FMNH P25112. Riggs pensó que los depósitos tenían similares en edad a las de la Como Bluff en Wyoming, de la que Marsh describió Brontosaurus. La mayor parte del esqueleto fue encontrado, y después de la comparación tanto con Brontosaurus y A. ajax , Riggs se dieron cuenta de que el holotipo de A. ajax era un ejemplar inmaduro, y por lo tanto las características que distinguen a los géneros no eran válidas. Ya que Apatosaurus fue nombrado con anterioridad, Brontosaurus debería considerarse como un sinónimo más moderno de Apatosaurus . Debido a esto, Riggs recombina B. excelsus como A. excelsus. Sobre la base de comparaciones con otras especies propuestas para Apatosaurus, Riggs también determinó que la muestra Museo Field Columbian fue probablemente lo más parecido a A. excelsus[12] A pesar de la publicación de Riggs, Henry Fairfield Osborn , que era un fuerte oponente de Marsh y su taxones, etiquetado el montaje de Apatosaurus del Museo Americano de Historia Natural Brontosaurius.[22][20] Debido a esta decisión el nombre Brontosaurus era de uso común fuera de la literatura científica para lo que Riggs considera Apatosaurus y la popularidad del museo significaba que Brontosaurus se convirtió en uno de los dinosaurios más conocidos, a pesar de que no era válido en toda casi la totalidad de los siglos 20 y principios del 21.[31]
Casi todos los paleontólogos modernos estuvieron de acuerdo con Riggs que Apatosaurus y Brontosaurus debían clasificarse juntos en un solo género. De acuerdo con las reglas del ICZN, que regula los nombres científicos de los animales, el nombre Apatosaurus, habiendo sido publicado por primero, tiene prioridad como el nombre oficial, Brontosaurus era considerado un sinónimo más moderno y por lo tanto fue largamente descartado del uso formal.[25][32][33][34] A pesar de esto, por lo menos un paleontólogo, Robert Bakker, sostuvo en los años 1990 que A. ajax y A. excelsus son de hecho suficientemente distintos para continuar mereciendo un género separado.[35] Esta idea sin embargo no contó con el apoyo de la mayor parte de la comunidad científica.[33][34]
En 2015 Emanuel Tschopp, Octávio Mateus y Roger Benson publicaron un artículo sobre la sistemática de los diplodócidos, y propusieron que los géneros podrían ser diagnosticados con base en 13 características diferentes, mientras que las especies se podían separar con 6. El número mínimo para la separación de los géneros fue escogido basándose en el hecho de que A. ajax y A. louisae difieren en 12 características, y Diplodocus carnegiei y D. hallorum difieren en 11 características. Por lo tanto, se escogieron 13 características para validar la separación de los géneros. Los 6 rasgos diferenciadores para la separación de especies se escogieron al contar el número de rasgos diferenciadores en especímenes separados que generalmente son considerados como pertenecientes a una sola especie, con solo un rasgo diferenciador en D. carnegiei y A. louisae, pero cinco rasgos en B. excelsus. Por lo tanto, Tschopp et al. afirmaron que Apatosaurus excelsus, clasificado originalmente como Brontosaurus excelsus, tenía suficientes diferencias morfológicas respecto a otras especies de Apatosaurus como para garantizar ser reclasificado de nuevo como un género distinto. La conclusión estaba basada en la comparación de 477 características morfológicas en 81 individuos diferentes de dinosaurios. Entre las muchas diferencias notables están el cuello más ancho -y presumiblemente más fuerte- de las especies de Apatosaurus comparado con el de B. excelsus. Otras especies previamente asignadas a Apatosaurus, tales como Elosaurus parvus y Eobrontosaurus yahnahpin fueron también reclasificadas como parte de Brontosaurus. Algunos de los rasgos propuestos para separar a Brontosaurus de Apatosaurus incluyen: vértebras dorsales posteriores con el centro más largo que ancho; la parte posterior de la escápula al borde del acromión y la extensión distal están excavadas; el borde del acromion de la extensión distal de la escápula tienen una expansión redondeada; y el radio de la longitud proximodistal a la anchura transversal del astrágalo es de 0.55 o mayor.[27] El paleontólogo Michael D'Emic hizo una crítica.[36] El paleontólogo Donald Prothero criticó la reacción de los medios de comunicación masivos de este estudio que consideró como superficial y prematuro, concluyendo :
Hasta que alguien resuelva forma convincente el tema, voy a poner "Brontosaurus" entre comillas y no seguir la última moda de los medios de comunicación, ni voy a hacer caso omiso de Riggs (1903) y poner el nombre en mis libros como un género válido.[37]
Clasificación
editarApatosaurus es un miembro de la familia Diplodocidae, un clado de dinosaurios saurópodos gigantescos. La familia incluye algunas de las criaturas más largas que jamás hayan caminado por la tierra, incluyendo Diplodocus, Supersaurus y Barosaurus. Apatosaurus es usualmente ubicado en su propia subfamilia, Apatosaurinae, junto con sus cercanos parientes Supersaurus, Eobrontosaurus y Brontosaurus.[38][39][40][13] Othniel Charles Marsh describió Apatosaurio como emparentado con Atlantosaurus dentro del grupo ya desaparecido Atlantosauridae.[12][26] En 1878, Marsh elevó a su familia a la categoría de suborden, incluyendo Apatosaurus, Atlantosaurus , Morosaurus (=Camarasaurus) y Diplodocus. Se clasifica dentro de este grupo Sauropoda, un grupo erigió en el mismo estudio.
En 1877, Othniel Charles Marsh publicó el nombre de la especie tipo Apatosaurus ajax. Continúa en 1879 con la descripción de otro espécimen más completo, que él pensó que representaba un nuevo género y lo llamó Brontosaurus excelsus. En 1903, Elmer Riggs argumentó que Brontosaurus excelsus era tan similar a Apatosaurus ajax que pertenece al mismo género, y por tanto lo reclasificó como Apatosaurus excelsus.[12] Desde que Riggs publicara sus opiniones, casi todos los paleontólogos han acordado que las dos especies deben ser clasificadas juntas en un solo género. De acuerdo con las reglas de ICZN el nombre Apatosaurus fue el primero en publicarse, dejando a Brontosaurus como sinónimo y descartándolo para su uso oficial. Brontosaurus es considerado como sinónimo más moderno y debería ser descartado en el uso formal.[32][41] En 2011, John Whitlock publicó un estudio que coloca Apatosaurus un diplodócido más basal pero menos basal de Supersaurus.[42][43]
Cladograma de la Diplodocidae según Tschopp, Mateus, y Benson (2015) donde se recupera Brontosaurus.[27]
| Diplodocidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Especies
editarMúltiples especies de Apatosaurus han sido designados a partir de material escaso. Marsh nombró muchas especies como pudo, lo que resultó en que muchos de ellos se basen en restos fragmentarios e indistinguibles. En 2005, Paul Upchurch et al. publicaron un estudio que analizó las especies y muestras de las relaciones de Apatosaurus. Encontraron que A. louisae fue la especie más basal, seguido de FMNH P25112, y luego una politomia de A. ajax , A. parvus , y A. excelsus.[16] Su análisis fue revisado y ampliado con muchos especímenes de diplodócido adicionales en 2015, que se resolvieron las relaciones de Apatosaurus de forma ligeramente diferente, y también apoyaron la separación Brontosaurus de Apatosaurus.[27]
Apatosaurus ajax
editarLa especie fue propuesta por Marsh en 1877 y su nombre procede de Ajax, un héroe de la mitología griega.[44] Marsh designa como holotipo el ejemplar siglado como YPM 1860, un esqueleto incompleto de un individuo juvenil. La especie está menos estudiada que Brontosaurus y A. louisae, sobre todo debido a la naturaleza incompleta del holotipo. En 2005, muchos especímenes, además del holotipo se encontraron asignable a A. ajax , YPM 1840, EMSN PV-20375, YPM 1861, y AMNH 460. Las muestras datan de finales del Kimeridgiano a los principios Titoniano.[16] En 2015, sólo el holotipo de A. ajax, YPM 1860, se asigna a la especie, con AMNH 460 encontrado ya sea para estar dentro de Brontosaurus, o potencialmente su propio taxón. Sin embargo, YPM 1861 y EMSN-PV 20375 solamente difieren en algunas características, y no se pueden distinguir específicamente o genéricamente de A. ajax. YPM 1861 es el holotipo de "Atlantosaurus" immanis, lo que significa que podría ser un sinónimo más moderno (1878) de A. ajax.[27]
Apatosaurus louisae
editarFue nombrado por Holland en 1916 haciendo referencia a Louise Carnegie, esposa de Andrew Carnegie quien financió la investigación de campo para encontrar los esqueletos completos del dinosaurio en el oeste americano. Siendo conocido por primera vez a partir de un esqueleto parcial que se encontró en Utah.[45] El holotipo es CM 3018, ca la que se asignaron los especímenes CM 3378, CM 11162 y LACM 52844. Los dos primeros consisten en una columna vertebral, los dos últimos consisten en un cráneo y un esqueleto casi completo, respectivamente. Sus ejemplares todos vienen del tarde Kimmeridgiano tardío del Monumento Nacional Dinosaurio.[16] En 2015, Tschopp et al. encontrado el espécimen tipo de A. laticollis anidado en estrecha cercanía con CM 3018, lo que significa que el primero es probablemente un sinónimo más moderno de A. louisae.[27] El cladograma a continuación es el resultado de un análisis realizado por Tschopp, Mateus, y Benson (2015). Los autores analizaron muchos especímenes tipo de diplodócidos por separado para deducir a la especie y género que pertenecían.[27]
| Apatosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Especies reasignadas
editar- Apatosaurio grandis fue nombrado en 1877 por Marsh en el artículo que describe A. ajax. Fue descrito brevemente, dibujado y diagnosticado.[3] Marsh mencionó más tarde que sólo fue asignado provisionalmente a Apatosaurus cuando se reasignó a su nuevo género Morosaurus en 1878.[46] Desde que Morosaurus ha sido considerado como un sinónimo de Camarasaurus, C. grandis es la especie más antigua nombrada de este último género.[47]
- Apatosaurio excelsus era la especie original tipo de Brontosaurus, nombrado por Marsh en 1879. Elmer Riggs reclasificado Brontosaurus como sinónimo de Apatosaurus en 1903, se transfirió B. excelsus a A. Excelsus . En 2015, Tschopp, Mateus, y Benson argumentaron que la especie era lo suficientemente clara para ser colocado en su propio género por lo que reclasificados de nuevo en Brontosaurus.[27]
- Apatosaurus parvus descrita por primera vez a partir de un ejemplar juvenil como Elosaurus en 1902 por Peterson y Gilmore, fue reasignado a Apatosaurio en 1994, y luego a Brontosaurus en 2015. Varias otras muestras, más maduros fueron asignados a él tras el estudio de 2015.[27]
- Apatosaurio minimus fue descrito originalmente como una muestra de Brontosaurus sp. en 1904 por Osborn. En 1917, Henry Mook nombró como su propia especie, A. minimus , para un par de huesos ilíacos y sacro.[3][48][49] En 2012, Mike P. Taylor y Matt J. Wedel publicó un breve resumen descriptivo del material de "A." minimous , encontrándolo difícil de ubicar entre Diplodocoidea y Macronaria. A pesar de que se colocó con Saltasaurus en un análisis filogenético, se cree que representan en su lugar alguna forma con características convergentes de muchos grupos.[49] El estudio de Tschopp et al. se encontró que una posición dentro de camarasáurido para el taxón fue apoyada, pero señaló se encontró que la posición del taxón ser muy variable y no había una posición más claramente probable.[27]
- Apatosaurio alenquerensis fue nombrado en 1957 por Albert-Félix de Lapparent y Georges Zbyweski. Se basó en el material craneal posterior de Portugal. En 1990, este material fue reasignado a Camarasaurus, pero en 1998 se le dio su propio género, Lourinhasaurus.[16] Esto fue apoyado además por los hallazgos de Tschopp et al. , en 2015, donde Lourinhasaurus se encontró que era hermano de Camarasaurus y otros camarasáuridos.[27]
- Apatosaurio yahnahpin fue nombrado por Filla James y Patrick Redman en 1994. Bakker hizo de A. yahnahpin la especie tipo de un género nuevo, Eobrontosaurus en 1998[35] y Tschopp lo reclasifico como Brontosaurio yahnahpin en 2015.[27]
Paleobiología
editarHasta la década de 1970 se pensó que los apatosaurios eran demasiado masivos para soportar su propio peso sobre terreno firme, por lo que se teorizó que estos saurópodos debían vivir en pantanos parcialmente sumergidos en el agua. Hallazgos más recientes no apoyan esta idea, ahora se piensa que los saurópodos han sido animales plenamente terrestres.[50] Los descubrimientos apoyaron la idea de que al igual que sus parientes los diplodocos, los apatosaurios vivían sobre suelo seco pastoreando, con su largo cuello y cola actuando como contrapesos. Un estudio ha descubierto que los diplodócidos tenían cuellos mucho menos flexibles que lo que se pensaba, no lo llevaban erguido como un cisne, sino en línea recta paralelo al suelo, de donde se alimentaban.[51] Un estudio sobre los hocicos de los diplodócido mostraron que el hocico cuadrado, los grandes pozos, y arañazos subparalelas finas de los dientes de los apatosaurio sugiere que realizaba un pastorea a baja altura no selectivo.[18] puede haber comido helechos, cycas, helechos con semilla, colas de caballo y algas.[51] Stevens y Parish en 2005 especularón que estos saurópodos se alimentaron desde orillas de los ríos de las plantas sumergidas en agua.[52]
Un estudio realizado en 2015, de los cuellos de apatosaurios y brontosaurio encontraron muchas diferencias entre ellos y otros diplodócidos, y que estas variaciones pueden haber demostrado que los cuellos de apatosaurios y brontosaurio fueron utilizados para el combate intraespecífico.[11] Varios usos para la única garra en la extremidad anterior de los saurópodos se han propuesto. Una sugerencia es que se utilizaron para la defensa, pero su forma y tamaño hace que esto sea improbable. También era posible que eran para la alimentación, pero el uso más probable de la garra era agarrar objetos tales como troncos de los árboles cuando eran juveniles.[14]
Rastros de saurópodos como apatosaurios muestran que ellos pueden haber tenido un rango de alrededor de 25-40 km por día, y que potencialmente podrían alcanzar una velocidades máximas de 20-30 km por hora.[10] La locomoción lenta de saurópodos puede ser debido a sus tamaños mínimos de musculatura, o para retroceder después de zancadas.[53] Estas pisadas demuestran que los apatosaurios jóvenes eran capaces de correr sobre sus patas traseras, asemejándose a un lagarto basilisco. En 2008 se publicaron los hallazgos,[54] aunque esto es discutido.[55][56] Las icnitas encontradas sugieren que vivían en manadas. En 2006 Matthew Mossbrucker descubrió huellas de un apatosaurio juvenil en la Quarry Five de Morrison, Colorado.
Postura del cuello
editarA principios del siglo XX, los diplodócidos, como los apatosaurios, eran retratados con sus largos cuellos erguidos, alimentándose de la vegetación de las copas de los árboles. Más recientemente, los científicos han sostenido que el corazón no habría podido crear la suficiente presión arterial para abastecer de oxígeno el cerebro.[57] Además, estudios más recientes han demostrado que la estructura de las vértebras del cuello no habría permitido que el cuello se doblara mucho hacia arriba y que saurópodos como Apatosaurus estaban adaptados al ramoneo bajo o a alimentarse del suelo.[51][52][58] Sin embargo, estudios posteriores demostraron que todos los tetrápodos parecen sostener el cuello a la máxima extensión vertical posible cuando está en una posición normal, en alerta, y sostiene que lo mismo sería válido para los saurópodos, sin conocer las características de la anatomía de los tejidos blandos del cuello de otros animales. Apatosaurus, como Diplodocus, habría elevado su cuello inclinado hacia arriba con la cabeza apuntando hacia abajo en una postura de descanso.[59][60] Kent Stevens y Michael Parrish en1999 y 2005 respectivamente afirman que los apatosaurios tenía una gran variedad de fuentes de alimentación. Su cuello podría doblarse en forma de U lateralmente.[51] El alcance del cuello del movimiento habría permitido también bajar la cabeza para alimentarse a la altura de los pies.[52]
En 2013, Mateo Cobley et al. entraron en esta controversia, encontrando que los grandes músculos y cartílagos permitirían un movimiento limitado del cuello. Afirman que los rangos de alimentación para los saurópodos como los diplodocos eran más pequeños de lo que se cree y que los animales pueden haber tenido que mover todo su cuerpo en búsqueda a mejores zonas de acceso de vegetación. Como tal, que podrían haber pasado más tiempo de forrajeo para satisfacer sus necesidades mínimas de energía.[61][62] Las conclusiones de Cobley et al. están en disputa con las de Taylor, quien analizó la cantidad y la colocación de cartílago intervertebrales para determinar la flexibilidad del cuello de los apatosaurios y diplodocos encontrando que el cuello de apatosaurio era muy flexible.[60]
Fisiología
editarDada la gran masa corporal y el cuello largo de los saurópodos como Apatosaurus, los fisiólogos han encontrado problemas en determinar cómo respiraban estos animales. Comenzando con la suposición de que al igual que los cocodrilos o las aves, Apatosaurus no tenía un diafragma, el volumen del volumen del espacio muerto , la cantidad de aire sin usar que quede en los tubos en la boca, la tráquea y el aire después de cada respiración, se ha estimado en alrededor de 184 litros por cada 30 toneladas de cuerpos. Paladino calcula su capacidad pulmonar, la cantidad de aire que se mueve en o fuera del cuerpo durante una sola respiración, sería de 904 l con un sistema respiratorio como las aves, 225 l como mamíferos, y 19 l como reptil.[63]
Sobre esta base, su sistema respiratorio no habría podido ser el propio de un reptil, pues su volumen de aire inspirado no habría podido sustituir el volumen en el espacio muerto. Asimismo, el sistema tipo mamífero proporcionaría solamente una fracción de aire nuevo en cada respiración. Su sistema respiratorio probablemente habría sido de parabronquios , con múltiples alvéolos pulmonares como en los pulmones aviares, y un pulmón de flujo a través. Un sistema respiratorio de las aves necesitaría un volumen pulmonar de aproximadamente 600 l en comparación con los de un mamífero de 2.950 l, lo que superaría el espacio disponible. El volumen torácico general de Apatosaurus se ha estimado en 1700 l, lo que permite un 500 l de un, corazón de cámaras cuatro y una capacidad pulmonar de 900 l. Que permitiría a aproximadamente 300 l para el tejido necesario.[63] La evidencia de un sistema respiratorio tipo aviar en Apatosaurius y otros saurópodos también está presente en la neumaticidad de las vértebras. Aunque esto juega un papel en la reducción del peso del animal, Wedel (2003) afirma que también es probable conectados a sacos de aire, como en las aves.[64]
James Spotila et al. en 1991 llega a la conclusión de que el gran tamaño corporal de los saurópodos les habría hecho incapaz de mantener altas tasas metabólicas, ya que no sería capaz de disipar suficiente calor.[65] Los autores asumieron que los saurópodos tenían un sistema respiratorio de reptil, Wedel dice que un sistema aviar lo habría permitido eliminar más calor.[64] Algunos científicos afirmar que el corazón habría tenido problemas para mantener una presión arterial suficiente para oxigenar el cerebro.[50] Otros sugieren que la postura casi horizontal de la de cabeza y cuello habría eliminado el problema de suministro de sangre al cerebro, ya que no habría que vencer la diferencia de altura.[64]
James Farlow en 1987 calcula que un Apatosaurus de cerca de 35 toneladas haya tenido 5,7 t de contenido de fermentación en su sistema digestivo.[66] Suponiendo Apatosaurus tenía un sistema respiratorio tipo ave y un metabolismo de reptil, Frank Paladino et al. (1997) estiman que el animal necesitaría consumir sólo alrededor de 262 litros de agua por día.[63] No se sabe cómo los apatosaurios podían comer suficiente alimento para satisfacer las necesidades de sus enormes cuerpos, pero es probable que comieran constantemente, deteniéndose brevemente para refrescarse, beber o quitarse parásitos. Se conjetura que dormían de pie. Confiaron probablemente en su tamaño y en el número de sus manadas para disuadir a los depredadores.
Patrones de crecimiento
editarUn estudio microscópico de 1999 de huesos de apatosaurios y brontosaurios concluyó los animales crecían rápidamente, de juveniles a tamaños casi adultos en unos 10 años.[67] En 2008, un estudio sobre las tasas de crecimiento de los saurópodos fue publicado por Thomas Lehman y Holly Woodward. Dijeron que mediante el uso de líneas de crecimiento y proporciones de longitud a la masa, un apatosaurio habría crecido hasta 25 toneladas en 15 años, con un crecimiento alcanzando un máximo de 5 t en un solo año. Un método alternativo, utilizando la longitud del miembro y la masa corporal, encontró que un apatosaurio creció 520 kg por año, y alcanzó su masa completa cerca de los 70 años de edad.[68] Estas estimaciones han sido consideradas poco fiable debido a los métodos de cálculo son no suena;buenos, las líneas de crecimiento más viejas habrían sido borradas por la remodelación ósea.[69] Uno de los primeros factores de crecimiento identificados de Apatosaurus era el número de vértebras sacras, que aumentó a cinco en el momento de la madurez de las criaturas. Esto fue observado por primera vez en 1903 y nuevamente en 1936.[3]
La histología de huesos largos permite a los investigadores estimar la edad que alcanza un individuo específico. Un estudio realizado por Eva Griebeler et al. en 2013 donde examinaron los datos histológicos de huesos largos y concluyó que el Apatosaurus sp. SMA 0014 pesaba 20.206 kg, alcanzó la madurez sexual a los 21 años, y murió 28. El mismo modelo de crecimiento de edad indicaron que el Apatosaurus sp. BYU 601 pesaba de 17.328 a 18,178 kg, alcanzando la madurez sexual a los 19 años, y murió a los 31.[69]
Juveniles
editarEn comparación con la mayoría de los saurópodos, una cantidad relativamente grande de material juvenil es conocido para Apatosaurus. Varios ejemplares en el Museo de Historia Natural de Oklahoma Sam Noble son de los juveniles de una especie indeterminada de Apatosaurus. Este material incluye un hombro parcial y la cintura pélvica, algunas vértebras y huesos de las extremidades. El material juvenil del OMNH es de al menos de dos diferentes grupos de edad y se basa en la superposición de huesos que probablemente proviene de más de tres individuos. Los ejemplares exhiben características que distinguen a Apatosaurus de sus relativos, y por lo tanto es probable que pertenecen al género.[16][70] Los saurópodos juveniles tienden a tener cuellos y colas proporcionalmente más cortos, y una disparidad extremidad anterior extremidad posterior más pronunciada que en los adultos.[71]
Cola
editarUn artículo de noviembre de 1997 en la revista Discover Magazine comenta la investigación de la mecánica de la cola de los apatosaurios, realizada por Nathan Myhrvold, un informático de Microsoft. Myhrvold realizó una simulación de computadora de la cola, que en diplodócidos como los apatosaurios tenían una estructura muy larga, con un ahusamiento que se asemejaba a un látigo. Este modelado de la computadora sugirió que los saurópodos eran capaces de producir con ella un sonido similar a un estallido de alrededor de 200 decibelios, comparable al volumen del sonido de un cañón.[72][73] Una patología ha sido identificado en la cola de un apatosaurio, causada por un defecto de crecimiento. Dos vértebras caudales se fusionan sin problemas a lo largo de toda la superficie de articulación del hueso, incluyendo los arcos de las espinas neurales. Este defecto podría haber sido causado por la falta o la inhibición de la sustancia que forma los discos o las articulaciones intervertebrales.[74] Se ha propuesto que los látigos podrían haber sido utilizados en combate, pero las colas de los diplodócidos eran bastante gráciles y delgadas en comparación con las de Shunosaurus y los mamenchisáuridos, y por lo tanto golpear a otro animal con la cola perjudicaría gravemente la cola en sí.[73] Posteriormente, Baron (2020) consideró que el uso de la cola como un látigo es improbable debido al potencial daño catastrófico a los músculos y el esqueleto que podría causar el agitar su grande y pesada cola a tales velocidades. En cambio, propone que las colas podrían haberse usado como un órgano táctil para mantenerse en contacto con los individuos situados a los lados o detrás en un grupo mientras migraban, lo cual hubiera aumentado la cohesión y permitiría la comunicación entre los individuos a la vez que limitaría el uso de actividades de comunicación más limitantes y energéticamente más costosas como detenerse a buscar a los individuos dispersos, girarse para vigilar a los individuos situados detrás, o comunicarse vocalmente.[75]
Paleoecología
editarLos fósiles de estos animales han sido encontrados en la Cantera Quarry, Wyoming, y en sitios de Colorado, Oklahoma y Utah, presente en zonas estratigráficas 2-6 de la formación Morrison. Esta es una secuencia superficial de sedimentos marinos superficiales y aluviales que, según la datación radiométrica, data de entre 156,3 millones de años en su base[76] y 146,8 millones de años en la parte superior,[77] colocándola ente el Oxfordiano tardío y las primeras etapas del Titoniano representando todo el kimergidiano durante el período Jurásico Tardío. Esta formación se interpreta como ambiente semiárido con estaciones secas y húmedas intercaladas. La cuenca Morrison, donde vivían los dinosaurios, se extendía desde Nuevo México hasta Alberta y Saskatchewan. Se formó cuando los precursores de la gama delantera de las montañas rocosas comenzó a empujar hacia el oeste. Los depósitos de sus cuencas de drenaje orientadas al Este se realizaron por arroyos y ríos y se depositan en las tierras bajas pantanosas, los lagos, los cauces fluviales y las llanuras de inundación.[78] Esta formación es similar en edad a la formación Lourinhã de Portugal y la formación Tendaguru en Tanzania.[79]
Apatosaurus es el segundo saurópodo más común en el ecosistema de la Formación Morrison después de Camarasaurus.[47] Apatosaurio puede haber sido más solitario que otros dinosaurios formación de Morrison.[80] Supersaurus tiene una mayor longitud total y es el más grande de todos los saurópodos de la formación Morrison.[13] Como género, Apatosaurus existió por un largo período de tiempo, y puede ser encontrado en muchos niveles de Morrison. Los fósiles de Apatosaurus ajax son conocidos exclusivamente de la porción superior de la formación, en el Miembro superior de la Cuenca Brushy, hace unos 152-151 millones de años. Los fósiles de A. excelsus se han encontrado en la parte superior del Miembro Salt Wash al Miembro superior de la Cuenca Brushy, que va desde mediados hasta finales del Kimmeridgiano, alrededor de 154-151 million años. Mientras que A. louisae presenta pocos fósiles, conocido solamente de un sitio del Miembro superior de la Cuenca Brushy, que data de la etapa Kimmeridgiano tardío, alrededor de 151 millones de años. Restos adicionales de Apatosaurus son conocidos a partir de rocas incluso más jóvenes, pero no han sido identificados como una especie en particular[81] y por lo tanto pueden pertenecer a Brontosaurus.[27]
La formación Morrison registra un momento en que el medio ambiente local estaba dominado por los dinosaurios saurópodos gigantescos.[47] Los dinosaurios conocidos de la Formación Morrison incluyen los terópodos Allosaurus, Ceratosaurus, Ornitholestes, Saurophaganax y Torvosaurus, los saurópodos Brontosaurus, Brachiosaurus, Camarasaurus y Diplodocus y los ornitisquios Camptosaurus, Dryosaurus y Stegosaurus.[82] Apatosaurus se encuentra comúnmente en los mismos sitios con Allosaurus, Camarasaurus, Diplodocus y Stegosaurus.[80] Allosaurus representó el 70-75% de las muestras de terópodos y estaba en la parte superior de la red trófica alimenticia de Morrison.[83] Muchos de los dinosaurios de la Formación Morrison son del mismo género que los observados en las rocas portuguesas de la formación lourinhã, principalmente Allosaurus, Ceratosaurus y Torvosaurus o tienen una contraparte cercana como Brachiosaurus y Lusotitan, Camptosaurus y Draconyx, Apatosaurus y Dinheirosaurus.[79] Otros vertebrados que se sabe que han compartido este paleoentorno se incluyen peces con aletas radiadas, ranas, salamandras , tortugas, esfenodontos, lagartos, terrestres y acuáticos cocodrilomorfos, y varias especies de pterosaurios . Conchas de los bivalvos y caracoles acuáticos también son comunes. La flora de la época se ha evidenciado en los fósiles de algas verdes, hongos, musgos, colas de caballo, cícadas, ginkgos y varias familias de coníferas. La vegetación variaba de bosques de galería en los ríos con helechos arborescentes y helechos, sabanas de helechos con árboles ocasionales como la araucaria Brachyphyllum.[84]
En la cultura popular
editarLos apatosaurios aparecen en la novela Parque Jurásico del escritor estadounidense Michael Crichton como una de las quince especies de dinosaurio recreados genéticamente, apareciendo también en la continuación de la novela El mundo perdido como una de las especies supervivientes en la isla Sorna o Enclave B.[85][86] En las películas basadas en las referidas novelas, los apatosaurios son reemplazados por ejemplares de Brachiosaurus en Parque Jurásico y Parque Jurásico III, y por Mamenchisaurus en El Mundo Perdido: Jurassic Park. En Jurassic World sí aparecen Apatosaurus como animales del parque.
Véase también
editarReferencias
editar- ↑ Holtz, Thomas (2012). Genus List for Holtz (2007) Dinosaurs:The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Consultado el 26 de septiembre de 2015.
- ↑ Foster, John (2007). Jurassic West: The Dinosaurs of the Morrison Formation and Their World (en inglés). Indiana: Indiana University Press. pp. 327-329. ISBN 978-0-253-34870-8.
- ↑ a b c d e f g h i j k l Gilmore, C.W. (1936). «Osteology of Apatosaurus, with special references to specimens in the Carnegie Museum». Memoirs of the Carnegie Museum 11 (4): 1-136. OCLC 16777126.
- ↑ Mazzetta, Gerardo V.; Christiansen, Per; Fariña, Richard A. (2004). «Giants and bizarres: body size of some southern South American Cretaceous dinosaurs» (PDF). Historical Biology: 1-13.
- ↑ Seebacher, Frank (2001). «A new method to calculate allometric length-mass relationships of dinosaurs». Journal of Vertebrate Paleontology 21 (1): 51-60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2. Consultado el 24 de febrero de 2016.
- ↑ Bates, Karl T.; Falkingham, Peter L.; Macaulay, Sophie; Brassey, Charlotte; Maidment, Susannah C. R. (2015). «Downsizing a giant: re-evaluating Dreadnoughtus body mass». Biology Letters (Royal Society Plubishing) (11): 1-6. ISSN 1744-957X.
- ↑ Wedel, Matt (2013). «A giant, skeletally immature individual of Apatosaurus from the Morrison Formation of Oklahoma». 61st Symposium on Vertebrate Palaeontology and Comparative Anatomy - Programme and Abstracts: 40-45. Consultado el 24 de febrero de 2016.
- ↑ Wedel, Matt (25 de abril de 2012). «The giant Oklahoma Apatosaurus: OMNH 1670». Sauropod Vertebra Picture of the Week. Consultado el 24 de febrero de 2016.
- ↑ Hartman, Scott (27 de abril de 2012). «Yup, the OK Apatosaurus is freakin' huge!». Scott Hartman's Skeletal Drawing.com. Consultado el 24 de febrero de 2016.
- ↑ a b c Fastovsky, D.E.; Weishampel, D.B. (2009). Dinosaurs: A Concise Natural History. Cambridge University Press. pp. 165-200. ISBN 978-0-521-88996-4.
- ↑ a b c Taylor, M.P.; Wedel, M.J.; Naish, Darren; Engh, B. (2015). «Were the necks of Apatosaurus and Brontosaurus adapted for combat?». PeerJ PrePrints 3: e1663. doi:10.7287/peerj.preprints.1347v1.
- ↑ a b c d e f g h Riggs, Elmer (1903). «Structure and Relationships of Opisthocoelian Dinosaurs. Part I, Apatosaurus Marsh». Publications of the Field Columbian Museum. Geological Series (2): 165-196.
- ↑ a b c d Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). «Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny». Arquivos do Museu Nacional 65 (4): 527-544. Archivado desde el original el 27 de julio de 2012.
- ↑ a b Upchurch, P. (1994). «Manus claw function in sauropod dinosaurs». Gaia 10: 161-171. ISSN 0871-5424.
- ↑ Martin, A.J. (2006). Introduction to the Study of Dinosaurs (Second ed.). Blackwell Publishing. p. 560. ISBN 1-4051-3413-5.
- ↑ a b c d e f Upchurch, P.; Tomida, Y.; Barrett, P.M. (2005). «A new specimen of Apatosaurus ajax (Sauropoda: Diplodocidae) from the Morrison Formation (Upper Jurassic) of Wyoming, USA». National Science Museum monographs 26 (118): 1-156. ISSN 1342-9574.
- ↑ a b c d McIntosh, J. S.; Berman, D.S. (1975). «Description of the palate and lower jaw of the sauropod dinosaur Diplodocus, (Reptilia: Saurischia) with remarks on the nature of the skull of Apatosaurus». Journal of Paleontology (Cambridge: Cambridge University Press) (49): 187-199. ISSN 0022-3360.
- ↑ a b c Whitlock, J.A. (2011). «Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses». PLoS ONE 6 (4): e18304. Bibcode:2011PLoSO...618304W. PMC 3071828. PMID 21494685. doi:10.1371/journal.pone.0018304.
- ↑ a b Balanoff, A.M.; Bever, G.S.; Ikejiri, T. (2010). «The Braincase of Apatosaurus (Dinosauria: Sauropoda) Based on Computed Tomography of a New Specimen with Comments on Variation and Evolution in Sauropod Neuroanatomy». American Museum Novitates 3677 (3677): 1-32. doi:10.1206/591.1.
- ↑ a b c d Parsons, K.M. (1997). «The Wrongheaded Dinosaur». Carnegie Magazine. Archivado desde el original el 14 de abril de 2010.
- ↑ Glut, D.F. (1997). Dinosaurs: The Encyclopedia. McFarland. pp. 150–163. ISBN 978-0-7864-7222-2.
- ↑ a b c d Miller, B. (30 de octubre de 2014). «Bully for Camarasaurus». Dinosours.
- ↑ Carpenter, Kenneth; Tidwell, Virginia (1998). «Preliminary Description of a Brachiosaurus Skull from Felch Quarry 1, Garden Park, Colorado». En Carpenter, Kenneth; Chure, Dan; Kirkland, James Ian, eds. The Upper Jurassic Morrison Formation: an interdisciplinary study. ISBN 978-90-5699-183-8.
- ↑ Barrett, P.M.; Storrs, G.W.; Young, M.T.; Witmer, L.M. (2011). «A new skull of Apatosaurus and its taxonomic and palaeobiological implications». Symposium of Vertebrate Palaeontology & Comparative Anatomy Abstracts of Presentations: 5.
- ↑ a b Taylor, M.P. (2010). «Sauropod dinosaur research: a historical review». Geological Society, London, Special Publications 343 (1): 361-386. Bibcode:2010GSLSP.343..361T. doi:10.1144/SP343.22.
- ↑ a b c Marsh, O.C. (1877). «Notice of New Dinosaurian Reptiles from the Jurassic formation». American Journal of Science 14 (84): 514-516.
- ↑ a b c d e f g h i j k l m n ñ Tschopp, E.; Mateus, O. V.; Benson, R. B. J. (2015). «A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)». PeerJ 3: e857. doi:10.7717/peerj.857.
- ↑ Holtz, T.R. Jr. (2008). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House. pp. 1–432. ISBN 0-375-82419-7.
- ↑ Marsh, O.C. (1879). «Notice of new Jurassic dinosaurs». American Journal of Science 18: 501-505.
- ↑ Bakker, Robert (1994). «The Bite of the Bronto». Earth 3 (6): 26-33. ISSN 1056-148X.
- ↑ Crafton, D.C. (1982). Before Mickey: The Animated Film 1898–1928. MIT Press. pp. 1-57. ISBN 0-262-03083-7.
- ↑ a b Brinkman, P. (2006). «Bully for Apatosaurus». Endeavour 30 (4): 126-130. PMID 17097734. doi:10.1016/j.endeavour.2006.10.004.
- ↑ a b Upchurch, P., Barrett, P.M. and Dodson, P. (2004). "Sauropoda". In Weishampel, D.B., Osmólska, H., and Dodson, P. (eds.), The Dinosauria (2nd edition). University of California Press, Berkeley 259-322.
- ↑ a b McIntosh, J.S. (1995). "Remarks on the North American sauropod Apatosaurus Marsh". Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers, A. Sun and Y. Wang (eds.), China Ocean Press, Beijing 119-123
- ↑ a b Bakker, R.T. (1998). "Dinosaur mid-life crisis: the Jurassic-Cretaceous transition in Wyoming and Colorado". In: S.G. Lucas, J.I. Kirkland, & J.W. Estep (eds.) Lower and Middle Cretaceous Terrestrial Ecosystems; New Mexico Museum of Natural History and Science Bulletin, 14: 67-77.
- ↑ D'Emic, M. 2015. "Not so fast, Brontosaurus". Time.com
- ↑ Prothero, D. 2015. "Is "Brontosaurus" Back? Not So Fast!". Skeptic.com.
- ↑ Lovelace, David M. (2007). «Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny». Arquivos do Museu Nacional 65 (4): 527-544.
- ↑ Taylor, M.P. and Naish, D. (2005). "The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda)." PaleoBios, 25(2): 1-7. (download here)
- ↑ Harris, J.D. (2006). "The significance of Suuwassea emiliae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution." Journal of Systematic Palaeontology, 4(2): 185–198.
- ↑ Taylor, M. P. (2010). "Sauropod dinosaur research: A historical review". Geological Society, London, Special Publications 343: 361. doi:10.1144/SP343.22. edit
- ↑ Whitlock, J.A. (2011). «A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda)». Zoological Journal of the Linnean Society 161 (4): 872-915. doi:10.1111/j.1096-3642.2010.00665.x.
- ↑ Gallina, P.A.; Apesteguía, S.; Haluza, A.; Canale, J.A. (2014). «A Diplodocid Sauropod Survivor from the Early Cretaceous of South America». En Farke, Andrew A., ed. PLoS ONE 9 (5): e97128. Bibcode:2014PLoSO...997128G. PMID 24828328. doi:10.1371/journal.pone.0097128.
- ↑ Carpenter, Kenneth; Currie, P.J., eds. (1992). Dinosaur Systematics, Approaches and Perspectives. Cambridge University Press. pp. 1–318. ISBN 0-521-36672-0.
- ↑ Carpenter, K.; McIntosh, J. (1994). «Upper Jurassic sauropod babies from the Morrison Formation». En Carpenter, Kenneth; Hirsch, Karl F.; Horner, John R., eds. Dinosaur Eggs and Babies. Cambridge University Press. pp. 265-278. ISBN 978-0-521-56723-7.
- ↑ Marsh, O.C. (1878). «Principal Characters of American Jurassic Dinosaurs». American Journal of Science 16 (95): 412-414.
- ↑ a b c Foster, John (2007). Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. p. 273. ISBN 978-0-253-34870-8.
- ↑ Taylor, M.P. (27 de julio de 2012). «"Apatosaurus" minimus sacrum/ilia, right lateral view». Sauropod Vertebrae Picture of the Week.
- ↑ a b Taylor, M.P.; Wedel, M.J. (2012). «Re-evaluating "Apatosaurus" minimus, a bizarre Morrison Formation sauropod with diplodocoid and macronarian features». SVPCA 2012 Programme and Abstracts: 23.
- ↑ a b Pierson, D.J. (2009). «The Physiology of Dinosaurs: Circulatory and Respiratory Function in the Largest Animals Ever to Walk the Earth». Respiratory Care 54 (7): 887-911. PMID 19558740. doi:10.4187/002013209793800286.
- ↑ a b c d Stevens, Kent A.; Parrish, J. Michael (1999). «Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs». Science 284 (5415): 798-800. doi:10.1126/science.284.5415.798. Consultado el 3 de agosto de 2008.
- ↑ a b c Stevens KA, Parrish JM (2005). «Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs». En Carpenter, Kenneth and Tidswell, Virginia (ed.), ed. Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 212–232. ISBN 0-253-34542-1. OCLC 218768170 57202057 61128849.
- ↑ Sellers, W.I.; Margetts, L.; Coria, R.A.; Manning, P.L. (2012). «March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs». PLoS ONE 8 (10): e78733. PMC 3864407. PMID 24348896. doi:10.1371/journal.pone.0078733.
- ↑ Rajewski, Genevieve (mayo de 2008). «Where Dinosaurs Roamed». Smithsonian: 20-24. Archivado desde el original el 15 de octubre de 2009. Consultado el 30 de abril de 2008.
- ↑ Switek, B. (2 de noviembre de 2010). «Did Wee Little Sauropods Stand Up to Run?». Smithsonian.com. Consultado el 20 de septiembre de 2015.
- ↑ «Tracks of a running bipedal baby brontosaur? Baby sauropod footprints discovered in Colorado». Science Daily (The Geological Society of America). 1 de noviembre de 2010. Consultado el 20 de septiembre de 2015.
- ↑ Pierson, D. J. (2009). "The Physiology of Dinosaurs: Circulatory and Respiratory Function in the Largest Animals Ever to Walk the Earth". Respiratory Care 54 (7): 887–911. doi:10.4187/002013209793800286. PMID 19558740. edit
- ↑ Upchurch, P, et al. (2000). «"Neck Posture of Sauropod Dinosaurs"» (PDF). Science 287, 547b (2000); DOI: 10.1126/science.287.5453.547b. Consultado el 5 de agosto de 2008.
- ↑ Taylor, M.P.; Wedel, M.J.; Naish, D. (2009). «Head and neck posture in sauropod dinosaurs inferred from extant animals». Acta Palaeontologica Polonica 54 (2): 213-220. doi:10.4202/app.2009.0007.
- ↑ a b Taylor, M.P. (2014). «Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs». PeerJ 2: e712. PMC 4277489. PMID 25551027. doi:10.7717/peerj.712.
- ↑ Cobley, M.J.; Rayfield, E.J.; Barrett, P.M. (2013). «Inter-Vertebral Flexibility of the Ostrich Neck: Implications for Estimating Sauropod Neck Flexibility». PLoS ONE 8 (8): e72187. doi:10.1371/journal.pone.0072187.
- ↑ Ghose, T. (15 de agosto de 2013). «Ouch! Long-Necked Dinosaurs Had Stiff Necks». livescience.com. Consultado el 31 de enero de 2015.
- ↑ a b c Paladino, F.V., Spotila, J.R., and Dodson, P. (1997). "A Blueprint for Giants: Modeling the Physiology of Large Dinosaurs." In Farlow, J.O. and Brett-Surman, M.K. (eds.), The Complete Dinosaur, Indiana University Press, 491-504. doi0253333490.
- ↑ a b c Wedel, M.J. (2003). «Vertebral Pneumaticity, Air Sacs, and the Physiology of Sauropod Dinosaurs». Paleobiology 29 (2): 243-255. JSTOR 4096832. doi:10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2.
- ↑ Spotila, J.R.; O'Connor, M.P.; Dodson, P.R.; Paladino, F.V. (1991). «Hot and cold running dinosaurs. Metabolism, body temperature, and migration». Modern Geology 16: 203-227.
- ↑ Farlow, J.A. (1987). «Speculations About the Diet and Physiology of Herbivorous Dinosaurs». Paleobiology 13 (1): 60-72. JSTOR 2400838.
- ↑ Curry, K.A. (1999). «Ontogenetic histology of Apatosaurus (Dinosauria: Sauropoda): new insights on growth rates and longevity». Journal of Vertebrate Paleontology 19 (4): 654-665. JSTOR 4524036. doi:10.1080/02724634.1999.10011179.
- ↑ Lehman, T.M.; Woodward, H.N. (2008). «Modelling growth rates for sauropod dinosaurs». Paleobiology 34 (2): 264-281. doi:10.1666/0094-8373(2008)034[0264:MGRFSD]2.0.CO;2.
- ↑ a b Griebeler, E.M.; Klein, N.; Sander, P.M. (2013). «Aging, Maturation and Growth of Sauropodomorph Dinosaurs as Deduced from Growth Curves Using Long Bone Histological Data: An Assessment of Methodological Constraints and Solutions». PLoS ONE 8 (6): e67012. doi:10.1371/journal.pone.0067012.
- ↑ Carpenter, K.; McIntosh, J.S. (1994). Carpenter, Kenneth; Hirsch, Karl. F.; Horner, John R., eds. Dinosaur Eggs and Babies. Cambridge University Press. pp. 265–274. ISBN 0-521-44342-3.
- ↑ Wedel, M. (2013). «Get down, get fuzzy, speculative juvenile Apatosaurus!». SVPOW.
- ↑ Zimmer, C. (1997). "Dinosaurs in Motion." Discover, 1 Nov 1997. [1] Accessed 27 Jul 2008.
- ↑ a b Myhrvold, N.P.; Currie, P.J. (1997). «Supersonic sauropods? Tail dynamics in the diplodocids». Paleobiology 23 (4): 393-409. JSTOR 2401127.
- ↑ Lovelace, D.M. (2014). «Developmental Failure of Segmentation in a Caudal Vertebra of Apatosaurus (Sauropoda)». The Anatomical Record 297 (7): 1262-1269. PMID 24532488. doi:10.1002/ar.22887.
- ↑ Baron, Matthew G. (3 de octubre de 2021). «Tactile tails: a new hypothesis for the function of the elongate tails of diplodocid sauropods». Historical Biology 33 (10): 2057-2066. ISSN 0891-2963. S2CID 219762797. doi:10.1080/08912963.2020.1769092.
- ↑ Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. (2006). «Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations». Geological Society of America Abstracts with Programs 38 (6): 7.
- ↑ Bilbey, S.A. (1998). «Cleveland-Lloyd Dinosaur Quarry - age, stratigraphy and depositional environments». En Carpenter, K.; Chure, D.; and Kirkland, J.I. (eds.), ed. The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. pp. 87-120. ISSN 0026-7775.
- ↑ Russell, D.A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. pp. 64-70. ISBN 978-1-55971-038-1.
- ↑ a b Mateus, O. (2006). «Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation 36. New Mexico Museum of Natural History and Science Bulletin. pp. 223-231.
- ↑ a b Dodson, Peter; Behrensmeyer, A.K.; Bakker, Robert T.; McIntosh, John S. (1980). «Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation». Paleobiology 6 (2): 208-232.
- ↑ Turner, C.E. and Peterson, F., (1999). "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A." Pp. 77–114 in Gillette, D.D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication 99-1.
- ↑ Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. (2006). «The fauna and flora of the Morrison Formation: 2006». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation 36. New Mexico Museum of Natural History and Science Bulletin. pp. 233-248.
- ↑ Foster, J.R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. 23. New Mexico Museum of Natural History and Science Bulletin. p. 29.
- ↑ Carpenter, K. (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus». En Foster, John R.; Lucas, Spencer G., eds. Paleontology and Geology of the Upper Jurassic Morrison Formation 36. New Mexico Museum of Natural History and Science Bulletin. pp. 131-138.
- ↑ Chrichton, Michael (1995) [1990]. Parque Jurásico (5 edición). Barcelona: Plaza & Janés. ISBN 84-01-49202-5.
- ↑ Chrichton, Michael (1997) [1995]. El Mundo Perdido (2 edición). Barcelona: Plaza & Janés. ISBN 84-01-49202-5.
Enlaces externos
editarPortal:Dinosaurios. Contenido relacionado con Dinosaurios.
Wikimedia Commons alberga una categoría multimedia sobre Apatosaurus.
Wikispecies tiene un artículo sobre Apatosaurus.
- Apatosaurus en duiops
- Apatosaurus
- www.dinolandia.com Información sobre dinosaurios
- Revista UNAM en-línea Tipos de Dinosaurios
- Apatosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Apatosaurus en la Dinosaur Encyclopaedia de Dino Russ's Lair (en inglés)
- Apatosaurus dentro de Diplodocoidea en The Thescelsosaurus (en inglés)
- Ficha e imagen de Apatosaurus en Dinosaurier-web (en alemán)
- Significado y pronunciación del nombre (en inglés)
Datos: Q14326