Glodari
| Glodari | |
|---|---|
 Vjeverica | |
| Sistematika | |
| Carstvo | Animalia |
| Koljeno | Chordata |
| Razred | Mammalia |
| Infraklasa | Eutheria |
| Nadred | Euarchontoglires |
| Red | Rodentia |
| Podredovi | |
Glodari (latinski: Rodentia) jesu najveći red sisara koje karakteriziraju po dva stalno rastuća sjekutića u gornjoj i donjoj vilici koja se konstantno troše.
U svijetu postoji približno 2.277 vrsta glodara, što čini gotovo 40% svih vrsta sisara,[1] a nastanjuju sve kontinente osim Antarktika. Glodari posjeduju oštre sjekutiće pomoću kojih sitne hranu, drvo ili grizu predatore. Većina glodara jede sjemenke ili biljke. Veličina tijela razlikuje se od vrste do vrste od približno 5 cm velikog afričkog patuljastog miša do približno 130 cm velikog vodenprasca, no većina vrsta velika je između 8 i 35 cm. Većina glodara noćne su životinje ili su aktivne u sumrak, ali se mogu sresti i po danu.
Vrste mogu da budu arborealne, fosorijalne (rijuće), ili semiakvatične. Neki od dobro poznatih glodara su: miševi, pacovi, vjeverice, dabrovi, hrčci, morsko prase i činčile. Druge životinje kao što su kunići, zečevi, i pikovi, čiji sjekutići isto tako konstantno rastu, su se nekada smatrali glodarima, ali se u današnje vrijeme smatraju zasebnim redom, Lagomorpha. Bez obzira na to, Rodentia i Lagomorpha su sestrinjske grupe, koje imaju najbližeg drevnog zajedničkog pretka i formiraju kladus Glires.
Većina glodara su male životinje sa robusnim tijelom, kratkim udovima i dugim repom. Oni koriste svoje oštre sjekutiće da glođu hranu, iskopavaju jazbine, i za samoodbranu. Većina glodara jede sjemenje ili drugi biljni materijal, dok neki od njih imaju raznovrsniju ishranu. Oni imaju tendenciju da budu društvene životinje i mnoge vrste žive u društvima sa kompleksnim vidovima međusobne komunikacije. Parenje između glodara može da varira od monogamije, do poliginije, do promiskuiteta. Mnogi imaju legla sa nerazvijenim mladuncima, dok su mladunci drugih predruštveni (relativno dobro razvijeni) pri rođenju.
Glodarski fosilni zapisi datiraju iz Paleocena na superkontinentu Laurazija. Raznovrsnost glodara je znatno porasla tokom Eocena, s njihovim širenjem po kontinentima, ponekad čak i prelazeći okeane. Glodari su dosegli Južnu Ameriku i Madagaskar iz Afrike, i oni su jedini kopneni placentalni sisari koji su dospjeli do i kolonizovali Australiju.
Glodari su korišteni kao hrana, za izradu odijela, kao kućni ljubimci, i kao laboratorijske životinje u istraživanjima. Neke vrste, a posebno smeđi pacov, crni pacov, i kućni miš, se smatraju štetočinama, koje jedu i upropaštavaju uskladištenu hranu, i šire bolesti. Slučajno uvedene vrste glodara se često smatraju invazivnim, i izazvale su izumiranje brojnih drugih vrsta, kao što su ostrvske ptice, prethodno izolovane od kopnenih predatora.
Karakteristike
[uredi | uredi izvor]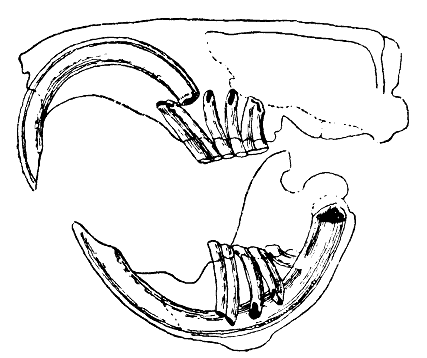

Prepoznatljiva karakteristika glodara su njihovi parovi sjekutića s otvorenim korijenom koji neprestano rastu [2]. Ovi sjekutići imaju debele slojeve cakline na prednjoj strani i malo cakline na stražnjoj strani [3]. Budući da ne prestaju da rastu, životinja ih mora i dalje istrošiti kako ne bi dosegnule i probile lubanju. Kako se sjekutići bruse jedan o drugi, mekši dentin na stražnjoj strani zuba se troši, ostavljajući oštru ivicu cakline u obliku oštrice dlijeta [4]. Većina vrsta ima do 22 zuba bez očnjaka ili prednjih pretkutnjaka. Razmak ili dijastema se javlja između sjekutića i obraznih zuba kod većine vrsta. Ovo omogućava glodavcima da usisavaju svoje obraze ili usne kako bi zaštitili svoja usta i grlo od strugotine i drugog nejestivog materijala, odbacujući ovaj otpad sa strane usta.
Kod mnogih vrsta, kutnjaci su relativno veliki i izbočeni. Kutnjaci glodara su dobro opremljeni za mljevenje hrane [5]. Muskulatura vilice je jaka. Donja vilica se gura naprijed tokom griženja i povlači se unazad tokom žvakanja [3]. Za griženje se koriste sjekutići, a za žvakanje kutnjaci, međutim, zbog kranijalne anatomije glodara ove metode hranjenja se ne mogu koristiti istovremeno i smatraju se međusobno isključivim [6]. Grupe glodara razlikuju se po rasporedu mišića čeljusti i povezanih struktura lobanje, kako od drugih sisara tako i međusobno. Sciuromorpha, kao što je istočna siva vjeverica, ima velik dubok masterični mišić, što je čini efikasnom u griženju sjekutićima. Myomorpha, kao što je smeđi štakor, ima uvećane sljepoočne i žvačne mišiće, što ih čini sposobnim da snažno žvaću svojim kutnjacima. Kod glodara, žvačni mišići se postavljaju iza očiju i doprinose pokretanju očiju koje se javlja tokom griženja, gdje brza kontrakcija i opuštanje mišića uzrokuje pomicanje očnih jabučica gore-dolje [7]. Hystricomorpha, kao što je zamorac, ima veće površne mišiće za žvakanje i manje duboke mišiće za žvakanje od štakora ili vjeverica, što ih možda čini manje efikasnim u griženju sjekutićima, ali njihovi uvećani unutrašnji pterygoidni mišići mogu im omogućiti da pomjeraju vilicu dalje u stranu [8]. Obrazna vrećica je specifična morfološka karakteristika koja se koristi za skladištenje hrane i evidentna je u određenim podgrupama glodara kao što su kengur pacovi, hrčci, vjeverice i gofovi koji imaju dvije vrećice koje se kreću od usta do prednjeg dijela ramena. Pravi miševi i pacovi nemaju ovu strukturu, ali su njihovi obrazi elastični zbog visokog stepena muskulature i inervacije u regiji [9].
Dok najveća vrsta, kapibara, može težiti čak 66 kg, većina glodara teži manje od 100 g. Glodavci obično imaju čučnjasta tijela i kratke udove. Prednji udovi obično imaju pet prstiju, uključujući suprotan palac, dok zadnji udovi imaju tri do pet prstiju. Lakat daje podlakticama veliku fleksibilnost. Većina vrsta je plantigradna, hodaju na dlanovima i tabanima i imaju nokte poput kandži. Nokti vrsta koje se ukopaju imaju tendenciju da budu dugi i jaki, dok glodari drveća imaju kraće i oštrije nokte. Različite vrste glodara koriste širok spektar metoda kretanja uključujući četveronožno hodanje, trčanje, kopanje, penjanje, dvonožno skakanje (klokani pacovi i miševi skakuću), plivanje, pa čak i klizanje. Ljuskaste vjeverice i leteće vjeverice, iako nisu u bliskom srodstvu, mogu kliziti s drveta na drvo koristeći membrane poput padobrana koje se protežu od prednjih do zadnjih udova [10]. Aguti ima brza stopala i nalik antilopi i nokte poput kopita. Većina glodara ima repove, koji mogu biti različitih oblika i veličina. Neki repovi služe kao hvataljke, npr. u evroazijskog žetvenog miša, a krzno na repovima može varirati od žbunastog do potpuno ćelavog. Rep se ponekad koristi za komunikaciju, kao kada dabrovi lupnu repom od površinu vode ili kućni miševi zveckaju repom da oglase uzbunu. Neke vrste imaju tek tragove repa ili ih uopšte nemaju. Kod nekih vrsta, rep je sposoban za regeneraciju ako se dio otkine.

Glodari uglavnom imaju dobro razvijena čula mirisa, sluha i vida. Noćne vrste često imaju uvećane oči, a neke su osjetljive na ultraljubičasto svjetlo. Mnoge vrste imaju dugačke, osjetljive brkove ili vibrize za dodir ili osjećaj vibracija iz okoline. Radnju brkova uglavnom pokreće moždano stablo, koje je samo provocirano korteksom. Međutim, Legg et al. 1989. pronalaze alternativni krug između korteksa i brkova kroz cerebelarne krugove, a Hemelt & Keller 2008. superiorni kolikulus [11]. Neki glodari imaju vrećice za obraze, koje mogu biti obložene krznom. One se mogu okrenuti naopačke radi čišćenja. Kod mnogih vrsta, jezik ne može doprijeti do sjekutića. Glodari imaju efikasan sistem za varenje, apsorbujući skoro 80% unesene energije. Kada jedete celulozu, hrana se omekšava u želucu i prenosi do cekuma, gdje je bakterije reduciraju na ugljikohidratne elemente. Glodavac tada prakticira koprofagiju, jedući vlastite fekalne pelete, tako da crijeva mogu apsorbirati hranjive tvari. Glodavci stoga često proizvode tvrdu i suhu fekalnu kuglicu. Horn et al. 2013.[12] navodi da glodari nemaju sposobnost povraćanja [13][14][15]. Kod mnogih vrsta penis sadrži kost, baculum; testisi mogu biti locirani ili abdominalno ili na preponama.
Spolni dimorfizam se javlja kod mnogih vrsta glodara. Kod nekih glodara mužjaci su veći od ženki, dok je kod drugih obrnuto. Seksualni dimorfizam pristranosti mužjaka tipičan je za vjeverice, kengure, i usamljene krtice; osobina koja se vjerovatno razvila zbog seksualne selekcije. Spolni dimorfizam pristranosti ženki postoji i među vjevericama i miševima koji skaču. Nije razumljivo zašto se ovaj obrazac pojavljuje, ali u slučaju nekih vrsta vjeverica, mužjaci su možda odabrali veće ženke zbog njihovog većeg reproduktivnog uspjeha. Kod nekih vrsta, kao što su voluharice, polni dimorfizam može varirati od populacije do populacije. Kod voluharica, ženke su tipično veće od mužjaka, ali seksualni dimorfizam u odnosu na mužjake javlja se u alpskim populacijama, vjerovatno zbog nedostatka predatora i veće konkurencije između mužjaka [16].
Rasprostranjenost i stanište
[uredi | uredi izvor]
Kao jedna od najrasprostranjenijih grupa sisara, glodari se mogu naći na svim kontinentima osim na Antarktiku. Oni su jedini kopneni placentarni sisari koji su kolonizirali Australiju i Novu Gvineju bez ljudske intervencije. Ljudi su također omogućili ovim životinjama da se šire na mnoga udaljena okeanska ostrva (npr. polinezijski pacov). Glodari su se prilagodili gotovo svakom kopnenom staništu, od hladne tundre (gdje mogu živjeti pod snijegom) do vrućih pustinja.
Neke vrste kao što su vjeverice na drvetu i dikobrazi Novog svijeta su arborealne, dok neke, poput gofova, tuco-tucosa i krtica, žive gotovo potpuno pod zemljom, gdje grade složene sisteme jazbina. Drugi žive na površini zemlje, ali mogu imati rupu u koju se mogu povući. Dabrovi i muskrati su poznati po tome što su poluvodeni, ali glodavac koji je najbolje prilagođen za život u vodi je vjerovatno vodeni pacov bez ušiju iz Nove Gvineje [17]. Glodari su također uspjeli živjeti u okolinama koje je stvorio čovjek, kao što su poljoprivredna i urbana područja [18].
Iako su neke vrste uobičajene štetočine za ljude, glodari također igraju važnu ekološku ulogu. Neki glodari se smatraju ključnim vrstama i inženjerima ekosistema u svojim staništima. U Velikim ravnicama Sjeverne Amerike, aktivnosti kopanja prerijskih pasa igraju važnu ulogu u aeraciji tla i preraspodjeli hranjivih tvari, podižući organski sadržaj tla i povećavajući apsorpciju vode. Oni održavaju ova staništa, a neki veliki biljojedi kao što su bizon i vilorog radije pasu u blizini kolonija prerijskih pasa zbog povećane nutritivne kvalitete stočne hrane [19].

Istrebljenje prerijskih pasa također može doprinijeti gubitku regionalnog i lokalnog biodiverziteta, povećanom uništavanju sjemena, te uspostavljanju i širenju invazivnog grmlja. Glodari koji se ukopaju mogu jesti plodna tijela gljiva i širiti spore kroz njihov izmet, dopuštajući na taj način gljivama da se rasprše i formiraju simbiotske odnose s korijenjem biljaka (koji obično ne mogu napredovati bez njih). Kao takvi, ovi glodari mogu igrati ulogu u održavanju zdravih šuma [20].
U mnogim umjerenim regijama, dabrovi igraju bitnu hidrološku ulogu. Kada grade svoje brane i kuće, dabrovi mijenjaju puteve potoka i rijeka [21] i omogućavaju stvaranje velikih močvarnih staništa. Jedna studija je pokazala da inženjering dabrova dovodi do 33 posto povećanja broja zeljastih biljnih vrsta u priobalnim područjima [22]. Druga studija je pokazala da dabrovi povećavaju populaciju divljeg lososa [23].
Ponašanje
[uredi | uredi izvor]Ishrana
[uredi | uredi izvor]
Većina glodara su biljojedi, hrane se isključivo biljnim materijalom kao što su sjemenke, stabljike, lišće, cvijeće i korijenje. Neki su svaštojedi, a neki su grabežljivci. Poljska voluharica je tipičan biljojed glodar i hrani se travama, biljem, korijenskim gomoljima, mahovinom i drugim rastinjem, a zimi grize koru. Povremeno jede beskičmenjake kao što su larve insekata [24]. Ravnični džepni goper jede biljni materijal pronađen pod zemljom tokom kopanja tunela, a također sakuplja travu, korijenje i gomolje u svojim obraznim vrećicama i čuva ih u podzemnim ostavima. Afrički pacov s vrećicom hrani se na površini, skupljajući sve što bi moglo biti jestivo u svoje prostrane obrazne vrećice dok mu lice ne izboči postrance. Zatim se vraća u svoju jazbinu kako bi sortirao materijal koji je prikupio i jede hranjive namirnice.
Agouti vrste su jedna od rijetkih grupa životinja koje mogu razbiti velike kapsule ploda brazilskog oraha. Unutra je previše sjemenki da bi se konzumiralo u jednom obroku, tako da agouti nešto odnese i spremi, što pomaže u raspršivanju sjemena. Ostala stabla koja rađaju orašaste plodove imaju tendenciju da obilato rađaju u jesen. Plodovi im budu previše brojni da bi se pojeli u jednom obroku, a vjeverice skupljaju i pohranjuju višak u pukotinama i šupljim stablima. Mnogo toga sjemenja kasnije proklija i izraste. U pustinjskim regijama sjeme je često dostupno samo na kratko. Kengur pacov skuplja sve što može pronaći i skladišti u svojoj jazbini [25].

Strategija za suočavanje sa sezonskim obiljem je da se jede što je više moguće i da se višak hranljivih sastojaka skladišti u obliku masti. Svizci to rade i mogu biti 50% teži u jesen nego u proljeće. Oslanjaju se na svoje rezerve masti tokom duge zimske hibernacije. Dabrovi se hrane lišćem, pupoljcima i unutrašnjom korom drveća koje raste, kao i vodenim biljkama. Hranu za zimu spremaju tako što u jesen obaraju mala stabla i lisnate grane i uranjaju ih u svoj ribnjak, zabadajući krajeve u blato kako bi ih usidrili. Ovdje mogu pristupiti zalihama hrane pod vodom čak i kada im je ribnjak zaleđen [26].
Iako se glodari tradicionalno smatraju biljojedima, većina malih glodavaca oportunistički uključuje insekte, crve, gljive, ribu ili meso u svoju prehranu, a neki su postali specijalizirani za ishranu životinjskog porijekla. Funkcionalno-morfološka studija sistema zuba glodara podržava ideju da su primitivni glodari bili svaštojedi, a ne biljojedi. Proučavanja literature pokazuju da su brojni pripadnici Sciuromorpha i Myomorpha, kao i nekoliko pripadnika Hystricomorpha, ili uključili životinjsku hranu u svoju prehranu ili su bili spremni da jedu takvu hranu kada se ponudi u zatočeništvu. Pregled sadržaja želuca sjevernoameričkog bijelonog miša, koji se inače smatra biljojedim, pokazao je 34% životinjske tvari.
Specijaliziraniji mesožderi uključuju rovčice na Filipinima, koji se hrane insektima i beskičmenjacima mekog tijela, i australskog vodenog štakora, koji proždire vodene insekte, ribe, rakove, dagnje, puževe, žabe, ptičja jaja i vodene ptice [27]. Miš skakavac iz sušnih krajeva Sjeverne Amerike hrani se insektima, škorpionima i drugim malim miševima, a samo mali dio njegove ishrane je biljni materijal. Ima zdepasto tijelo sa kratkim nogama i repom, ali je okretan i lako može savladati plijen veliki poput njega samog.
Društveno ponašanje
[uredi | uredi izvor]
Glodari pokazuju širok raspon društvenih ponašanja od kastinskog sistema sisara golog krtičnjaka [28], opsežnog "grada" kolonijalnog prerijskog psa [29], preko porodičnih grupa do nezavisnog, usamljenog života puha. Odrasli puhovi mogu imati preklapajući raspon hranjenja, ali žive u pojedinačnim gnijezdima i hrane se odvojeno, okupljajući se nakratko u sezoni parenja. Džepni gof je također usamljena životinja izvan sezone parenja, svaka jedinka kopa složen sistem tunela i održava teritoriju.
Veći glodari obično žive u porodičnim jedinicama u kojima roditelji i njihovi potomci žive zajedno dok se mladi ne raziđu. Dabrovi žive u proširenim porodičnim jedinicama obično sa parom odraslih, ovogodišnjim i prošlogodišnjim potomcima, a ponekad i starijim mladima [30]. Smeđi štakori obično žive u malim kolonijama sa do šest ženki koje dijele jazbinu i jednim mužjakom koji brani teritoriju oko jazbine. Pri velikoj gustini naseljenosti, ovaj sistem se raspada i mužjaci pokazuju hijerarhijski sistem dominacije sa preklapajućim rasponima. Žensko potomstvo ostaje u koloniji dok se muški mladi raspršuju. Prerijska voluharica je monogamna i stvara doživotnu parnu vezu. Izvan sezone parenja, prerijske voluharice žive s drugima u malim kolonijama. Mužjak nije agresivan prema drugim mužjacima dok se ne pari, ali nakon parenja brani teritoriju, ženku i gnijezdo od drugih mužjaka. Par se okuplja, njeguje jedno drugo i dijeli odgovornosti za gniježđenje i uzgoj mladih [31].
Među najdruštvenijim glodarima su vjeverice, koje obično formiraju kolonije zasnovane na ženskom srodstvu, a mužjaci se raspršuju nakon odbijanja i postaju nomadski kao odrasli. Saradnja kod vjeverica varira među vrstama i obično uključuje alarmiranje, odbranu teritorije, dijeljenje hrane, zaštitu područja gniježđenja i sprječavanje čedomorstva [32]. Crnorepi prerijski pas formira velike gradove koji mogu pokrivati mnogo hektara. Jame nisu međusobno povezane, već su iskopane i zauzete od strane teritorijalnih porodičnih grupa poznatih kao koterije. Koteriju često čine odrasli mužjak, tri ili četiri odrasle ženke, nekoliko jednogodišnjaka koji se ne razmnožavaju i potomci tekuće godine. Pojedinci unutar koterija su prijateljski nastrojeni jedni prema drugima, ali neprijateljski prema strancima.
Možda najekstremniji primjeri kolonijalnog ponašanja kod glodara je kod slijepog kučeta. Oni žive potpuno pod zemljom i mogu formirati kolonije do 80 jedinki. U koloniji se razmnožavaju samo jedna ženka i do tri mužjaka, dok su ostali članovi manji i sterilni i rade kao radnici. Neki pojedinci su srednje veličine. Pomažu u odgoju mladih i mogu zamijeniti reprodukciju ako netko umre.
Komunikacija
[uredi | uredi izvor]Čulom mirisa
[uredi | uredi izvor]
Glodari koriste označavanje mirisa u mnogim društvenim kontekstima uključujući komunikaciju među i unutar vrsta, obilježavanje staza i uspostavljanje teritorija. Njihov urin pruža genetske informacije o pojedincima, uključujući vrstu, spol i individualni identitet, te metaboličke informacije o dominaciji, reproduktivnom statusu i zdravlju. Jedinjenja izvedena iz glavnog kompleksa histokompatibilnosti (MHC) vezana su za nekoliko proteina u urinu. Miris grabežljivca smanjuje učestalost označavanja mirisa.[45]
Glodari su u stanju prepoznati bliske srodnike po mirisu i to im omogućava da pokažu preferencijalno ponašanje prema svojim njima i izbjegavaju parenje između bližih srodnika. Ovo srodničko prepoznavanje se odvija pomoću mirisnih znakova iz urina, fecesa i sekreta žlijezda. Glavna procjena može uključivati MHC, gdje je stepen srodstva dvije osobe u korelaciji sa MHC genima koji su im zajednički. U komunikaciji bez srodstva, gdje su potrebni trajniji markeri mirisa, kao na teritorijalnim granicama, tada se mogu koristiti i nepromjenjivi glavni urinarni proteini (MUP), koji funkcionišu kao transporteri feromona. MUP-ovi također mogu signalizirati individualni identitet, pri čemu svaki mužjak kućnog miša (Mus musculus) izlučuje urin koji sadrži desetak genetski kodiranih MUP-ova [33].
Kućni miševi talože urin, koji sadrži feromone, za teritorijalno obilježavanje, individualno i grupno prepoznavanje i društvenu organizaciju. Teritorijalni dabrovi i crvene vjeverice istražuju i upoznaju se s mirisima svojih susjeda i manje agresivno reagiraju na njihove upade nego na one koje čine neteritorijalne "skitnice" ili stranci. Ovo je poznato kao "efekat dragog neprijatelja" [34].
Slušna
[uredi | uredi izvor]
Mnoge vrste glodara, posebno oni koji su dnevni i društveni, imaju širok spektar poziva alarma koji se emituju kada primijete prijetnje. Postoje i direktne i indirektne koristi od ovoga. Potencijalni grabežljivac se može zaustaviti kada zna da je otkriven, ili poziv alarma može omogućiti bližnjima da preduzmu akciju izbjegavanja [35]. Nekoliko vrsta, na primjer prerijski psi, imaju složene alarmne sisteme protiv predatora. Ove vrste mogu imati različite pozive za različite grabežljivce (npr. zračne grabežljivce ili grabežljivce na zemlji) i svaki poziv sadrži precizne informacije o prirodi prijetnje. Hitnost prijetnje također se prenosi akustičnim svojstvima poziva [36].
Društveni glodari imaju širi raspon vokalizacije od usamljenih vrsta. Petnaest različitih tipova poziva je prepoznato kod odraslih kataba krtica i četiri kod juvenila. Slično tome, obični degu, još jedan društveni glodar koji buši, pokazuje široku lepezu komunikacijskih metoda i ima razrađen vokalni repertoar koji se sastoji od petnaest različitih kategorija zvuka [37]. Ultrazvučni pozivi igraju ulogu u društvenoj komunikaciji između puhova i koriste se kada su jedinke van vidokruga jedni drugih.
Kućni miševi koriste i zvučne i ultrazvučne pozive u različitim kontekstima. Zvučne vokalizacije se često mogu čuti tokom agonističkih ili agresivnih susreta, dok se ultrazvuk koristi u seksualnoj komunikaciji, kao i kod mladih kada ispadnu iz gnijezda.
Vizuelna
[uredi | uredi izvor]Glodari, kao i svi placentni sisari osim primata, imaju samo dva tipa čunjića koji primaju svjetlost u svojoj mrežnjači [38], kratkotalasnu "plavo-UV" tip i srednje talasne dužine "zeleni" tip. Stoga su klasifikovani kao dihromati; međutim, oni su vizualno osjetljivi na ultraljubičasti (UV) spektar i stoga mogu vidjeti svjetlost koju ljudi ne mogu. Funkcije ove UV osjetljivosti nisu uvijek jasne. Kod degua, na primjer, trbuh reflektira više UV svjetla nego leđa. Stoga, kada degu ustane na stražnje noge, što čini kada je uzbunjen, izlaže svoj trbuh drugim deguima i ultraljubičasti vid može poslužiti u komunikaciji alarma. Kada stoji na sve četiri, njegova niska UV refleksija leđa bi mogla pomoći da degu bude manje vidljiv grabežljivcima. Ultraljubičastog svjetla ima u izobilju tokom dana, ali ne i noću. Postoji veliki porast omjera ultraljubičastog i vidljivog svjetla u jutarnjim i večernjim satima sumraka. Mnogi glodari su aktivni tokom sumraka (krepuskularna aktivnost), a UV osjetljivost bi bila korisna u tim vremenima. Ultraljubičasta refleksija je sumnjive vrijednosti za noćne glodare.
Urin mnogih glodara (npr. voluharice, degui, miševi, pacovi) snažno reflektuje UV svjetlo i to se može koristiti u komunikaciji ostavljajući vidljive, ali i mirisne oznake [39]. Međutim, količina UV koja se reflektuje opada s vremenom, što u nekim okolnostima može biti nepovoljno; vjetruška može razlikovati stare i svježe staze glodara i ima veći uspjeh u lovu na nedavno označenim rutama [40].
Dodirna
[uredi | uredi izvor]
Vibracije mogu dati naznake bližnjima o specifičnim ponašanjima, upozorenju i izbjegavanju predatora, održavanju stada ili grupe i udvaranju. Bliskoistočno slijepo kuče bilo je prvi sisavac za kojeg je dokumentirana seizmička komunikacija. Ovi glodari udaraju glavom o zidove svojih tunela. Ovo ponašanje je u početku tumačeno kao dio njihovog ponašanja u izgradnji tunela, ali se na kraju shvatilo da oni generiraju vremenski uzorkovane seizmičke signale za komunikaciju na daljinu sa susjednim krticama [41].
Bubnjanje se naširoko koristi kao upozorenje predatora ili odbrambena akcija. Koriste ga prvenstveno fosorni ili polufosorni glodari [42]. Klokanski štakor proizvodi nekoliko složenih obrazaca bubnjanja stopala u brojnim različitim kontekstima, od kojih je jedan kada naiđe na zmiju. Bubanj može upozoriti mladunce, ali najvjerovatnije govori da je pacov previše oprezan za uspješan napad, čime se sprječava zmijska grabežljiva potjera [43]. Nekoliko studija je ukazalo na namjernu upotrebu vibracija tla kao sredstva intraspecifične komunikacije tokom udvaranja među krticama Cape [44]. Prijavljeno je da je bubnjanje uključeno u muško-muško takmičenje; dominantni mužjak pokazuje svoj potencijal zaštite teritorije bubnjanjem, čime se minimizira fizički kontakt sa potencijalnim rivalima.
Strategije parenja
[uredi | uredi izvor]
Neke vrste glodara su monogamne, a odrasli mužjak i ženka stvaraju trajnu vezu u paru. Monogamija može doći u dva oblika; obavezna i fakultativna. U obaveznoj monogamiji, oba roditelja brinu o potomstvu i igraju važnu ulogu u njihovom opstanku. Ovo se dešava kod vrsta kao što su kalifornijski miševi, oldfield miševi, malagaški divovski pacovi i dabrovi. Kod ovih vrsta mužjaci se obično pare samo sa svojim partnericama. Pored povećane brige o mladima, obavezna monogamija može biti korisna i odraslom mužjaku jer smanjuje šanse da nikada ne nađe partnera ili da se pari sa neplodnom ženkom. U fakultativnoj monogamiji, mužjaci ne pružaju direktnu roditeljsku brigu i ostaju s jednom ženkom jer ne mogu pristupiti drugima zbog prostorne disperzije. Čini se da su prerijske voluharice primjer ovog oblika monogamije, gdje mužjaci čuvaju i brane ženke u njihovoj blizini [45].
Kod poliginih vrsta, mužjaci će pokušati monopolizirati i pariti se s više ženki. Kao i kod monogamije, poliginija kod glodara može doći u dva oblika; odbrambena i neodbrambena. Odbrambena poliginija uključuje muškarce koji kontroliraju teritorije koje sadrže resurse koji privlače ženke. To se događa kod vjeverica poput žutoglavih svizaca, kalifornijskih vjeverica, kolumbijskih vjeverica i Richardsonovih vjeverica. Mužjaci sa teritorijama poznati su kao "rezidentni" mužjaci, a ženke koje žive unutar teritorija poznate su kao "rezidentne" ženke. U slučaju svizaca, čini se da domaći mužjaci nikada ne gube svoju teritoriju i uvijek pobjeđuju u susretima s invazivnim mužjacima. Poznato je i da neke vrste direktno brane svoje ženke i borbe koje se dešavaju mogu dovesti do teških ranjavanja. Kod vrsta sa poliginijom bez odbrane, mužjaci nisu teritorijalni i lutaju naširoko u potrazi za ženkama. Oni uspostavljaju hijerarhiju dominacije, pri čemu visokorangirani mužjaci imaju pristup većini ženki. Ovo se dešava kod vrsta kao što su Beldingove vjeverice i neke vrste vjeverica na drvetu.

Majke glodari pružaju i direktnu roditeljsku brigu, kao što je dojenje, njegovanje, vraćanje i okupljanje, i indirektno roditeljstvo, kao što je čuvanje hrane, izgradnja gnijezda i zaštita njihovog potomstva [46]. U mnogim društvenim vrstama, o mladima se mogu brinuti pojedinci koji nisu njihovi roditelji, što je praksa poznata kao aloparenting ili kooperativni uzgoj. Poznato je da se to događa kod crnorepih prerijskih pasa i Beldingovih kopnenih vjeverica, gdje majke imaju zajednička gnijezda i doje nesrodne mlade zajedno sa svojima. Postavlja se pitanje da li ove majke mogu razlikovati koji su njihovi mladi. U patagonskih mara, mladi se također stavljaju u zajedničke jame (gnijezda), ali majke ne dozvoljavaju da ih doje druga mladunčad osim njihovih [47].
Čedomorstvo postoji kod brojnih vrsta glodara i mogu ga prakticirati odrasli srodnici oba spola. Predloženo je nekoliko razloga za ovakvo ponašanje, uključujući nutritivni stres, nadmetanje u resursima, izbjegavanje pogrešnog usmjeravanja roditeljske brige i, u slučaju muškaraca, pokušaj da se majka učini seksualno prijemčivom. Potonji razlog je dosta uočen kod primata i lavova, ali manje kod glodara [48]. Čini se da je čedomorstvo široko rasprostranjeno kod crnorepih prerijskih pasa, uključujući čedomorstvo od invazivnih mužjaka i ženki imigranata, kao i povremeni kanibalizam vlastitog potomstva pojedinca. Da bi se zaštitile od čedomorstva drugih odraslih , ženke glodara mogu koristiti izbjegavanje ili direktnu agresiju na potencijalne počinioce, višestruko parenje, teritorijalnost ili prijevremeni prekid trudnoće. Feticid se također može pojaviti među glodarima; kod alpskih svizaca dominantne ženke imaju tendenciju da potisnu reprodukciju podređenih tako što su antagonističke prema njima dok su trudne. Rezultirajući stres uzrokuje pobačaj fetusa [49].
Rađanje i roditeljstvo
[uredi | uredi izvor]
Glodari se mogu roditi ili altricijski (slijepi, bez dlake i relativno nerazvijeni) ili prekocijalni (uglavnom krzneni, otvorenih očiju i prilično razvijeni), u zavisnosti od vrste. Altricijalno stanje je tipično za vjeverice i miševe, dok se prekocijalno stanje obično javlja kod vrsta kao što su zamorci i dikobrazi. Ženke s altricijskim mladuncima obično grade složena gnijezda prije nego što se okote i održavaju ih dok se njihovo potomstvo ne osamostali. Ženka rađa sjedeći ili ležeći. Novorođenčad prvi put izađu iz gnijezda nekoliko dana nakon što su otvorili oči i u početku se redovno vraćaju. Kako su stariji i razvijeniji, rjeđe posjećuju gnijezdo i napuštaju ga za stalno obično kada se odbiju od dojenja [50].
Kod prekocijalnih vrsta, majke malo ulažu u izgradnju gnijezda, a neke uopće ne grade gnijezda. Ženka rađa stojeći. Majke ovih vrsta održavaju kontakt sa svojim visoko pokretnim mladuncima uz pomoć majčinih kontaktnih poziva. Iako su relativno nezavisni i odviknuti od dojenja u roku od nekoliko dana, ipak ih majke još neko vrijeme mogu nastaviti dojiti i njegovati. Veličine legla glodara također varira i ženke s manjim leglom provode više vremena u gnijezdu od onih s većim leglom.
Inteligencija
[uredi | uredi izvor]
Glodari imaju napredne kognitivne sposobnosti. Oni mogu brzo naučiti izbjegavati otrovane mamce, što ih čini teškim protivnicima prilikom borbe protiv štetočina.[Zamorci mogu naučiti i zapamtiti složene puteve do hrane. Vjeverice i kengur-pacovi su u stanju da lociraju zalihe hrane prostornom memorijom, a ne samo mirisom [51].
S obzirom da se laboratorijski miševi (kućni miševi) i pacovi (smeđi pacovi) naširoko koriste kao naučni modeli za dalje razumijevanje biologije, dostupna su mnoga saznanja o njihovim kognitivnim kapacitetima. Smeđi pacovi pokazuju kognitivnu pristrasnost, gdje je obrada informacija pristrasna ovisno o tome da li su u pozitivnom ili negativnom afektivnom stanju [52]. Na primjer, laboratorijski štakori obučeni da reagiraju na određeni ton pritiskom na polugu za primanje nagrade i da pritisnu drugu polugu kao odgovor na drugačiji ton kako bi izbjegli strujni udar, vjerojatnije će odgovoriti na srednji ton odabirom poluge nagrađivanja ako su ih upravo golicali (nešto u čemu uživaju), što ukazuje na "vezu između direktno izmjerenog pozitivnog afektivnog stanja i donošenja odluka pod neizvjesnošću u životinjskom modelu."
Laboratorijski (smeđi) štakori mogu imati sposobnost metakognicije – da razmotre svoje vlastito učenje, a zatim donose odluke na osnovu onoga što znaju ili ne znaju, kao što pokazuju izbori koje donose, očigledno mijenjajući težinu zadataka i očekivane nagrade, čineći ih prve životinje osim primata za koje se zna da imaju ovaj kapacitet [53], ali ovi nalazi su sporni, budući da su pacovi možda slijedili jednostavne principe operantnog kondicioniranja, ili ekonomski model ponašanja [54]. Smeđi pacovi koriste društveno učenje u širokom spektru situacija, ali možda posebno u sticanju preferencija u hrani.
Klasifikacija i evolucija
[uredi | uredi izvor]Evoluciona historija
[uredi | uredi izvor]
Zubi su ključna karakteristika po kojoj se fosilni glodari prepoznaju, a najraniji zapis o takvim sisarima potiče iz paleocena, ubrzo nakon izumiranja neptičjih dinosaurusa prije nekih 66 miliona godina. Ovi fosili se nalaze u Lauraziji [55], superkontinentu koji se sastoji od današnje Sjeverne Amerike, Evrope i Azije. Divergencija Gliresa, grupe koja se sastoji od glodara i lagomorfa (zečeva i pika), od drugih placentnih sisara dogodila se u roku od nekoliko miliona godina krede i prije paleogena. Neki podaci o molekularnom satu ukazuju na to da su se moderni glodari (pripadnici reda glodara) pojavili u kasnoj kredi, iako su druge procjene molekularne divergencije u skladu sa fosilnim zapisom [56].
Smatra se da su glodari evoluirali u Aziji, gdje su lokalne faune bile ozbiljno pogođene izumiranjem iz krede i paleogena i nikada se nisu u potpunosti oporavile, za razliku od njihovih sjevernoameričkih i evropskih rođaka. U nastalom ekološkom vakuumu, glodari su mogli evoluirati i diverzificirati se, zauzimajući upražnjene niše.
Hstorija kolonizacije svjetskih kontinenata od strane glodara je složena. Kretanja velike nadporodice Muroidea (uključujući hrčke, gerbile, prave miševe i pacove) su možda uključivala do sedam kolonizacija Afrike, pet Sjeverne Amerike, četiri Jugoistočne Azije, dvije Južne Amerike i do deset Evroazije [57].

Tokom eocena, glodari su počeli da se diverzifikuju. Dabrovi su se pojavili u Evroaziji u kasnom eocenu pre nego što su se proširili na Sjevernu Ameriku u kasnom miocenu. Kasno u eocenu, histrikognati se šire na Afriku, najvjerovatnije su nastali u Aziji prije najmanje 39,5 miliona godina. Iz Afrike, fosilni dokazi pokazuju da su neki histrikognati (kaviomorfi) kolonizirali Južnu Ameriku, koja je u to vrijeme bila izolirani kontinent, očito koristeći okeanske struje da pređu Atlantik na plutajućim krhotinama. Kaviomorfi su stigli u Južnu Ameriku prije 41 milion godina (što implicira da je datum barem isti za histrikognate u Africi) [58], i stigli su do Velikih Antila u ranom oligocenu, što sugerira da su se sigurno brzo raspršili po jugu Amerika.
Smatra se da su glodari Nesomyid splavarili iz Afrike na Madagaskar prije 20-24 miliona godina. Čini se da su svih 27 vrsta autohtonih malagaških glodara potomci jednog događaja kolonizacije.
Prije 20 miliona godina pojavili su se fosili koji su prepoznatljivo pripadali sadašnjim porodicama kao što su Muridae. Do miocena, kada se Afrika sudarila sa Azijom, afrički glodari poput dikobraza počeli su da se šire u Evroaziju. Neke fosilne vrste bile su veoma velike u poređenju sa modernim glodarima i uključivale su džinovskog dabra, Castoroides ohioensis, koji je narastao do dužine od 2,5 m i težine od 100 kg. Najveći poznati glodar bio je Josephoartigasia monesi, pacarana s procijenjenom dužinom tijela od 3 m.
Prvi glodari stigli su u Australiju preko Indonezije prije oko 5 miliona godina. Iako su tobolčari najistaknutiji sisari u Australiji, mnogi glodari, svi koji pripadaju potporodici Murinae, spadaju među vrste sisara na kontinentu. Postoji oko pedeset vrsta 'starih endema', prvi val glodara koji je kolonizirao zemlju u miocenu i ranom pliocenu, i osam pravih vrsta pacova (Rattus) 'novih endema', koji su stigli u sljedećem valu u kasnom pliocenu. ili ranog pleistocena. Najraniji fosilni glodari u Australiji imaju maksimalnu starost od 4,5 miliona godina, a molekularni podaci su u skladu sa kolonizacijom Nove Gvineje sa zapada tokom kasnog miocena ili ranog pliocena, nakon čega je uslijedila brza diverzifikacija. Dalji val adaptivne radijacije dogodio se nakon jedne ili više kolonizacija Australije nekih 2 do 3 miliona godina kasnije [59].
Glodari su učestvovali u Velikoj američkoj razmjeni koja je nastala spajanjem Amerike formiranjem Panamske prevlake, prije oko 3 miliona godina u doba Piacenziana. U ovoj razmjeni, mali broj vrsta kao što su dikobrazi Novog svijeta (Erethizontidae) krenuo je na sjever. Međutim, glavna invazija sigmodontina prema jugu prethodila je formiranju kopnenog mosta za najmanje nekoliko miliona godina, vjerovatno koristeći plutajuća debla kao splav [60]. Sigmodontinci su se jednom eksplozivno diverzificirali u Južnoj Americi, iako se određeni stepen diverzifikacije možda već dogodio u Centralnoj Americi prije kolonizacije.
Standardna klasifikacija
[uredi | uredi izvor]Upotreba naziva reda "Rodentia" pripisuje se engleskom putniku i prirodoslovcu Thomasu Edwardu Bowdichu (1821). Moderna latinska riječ Rodentia je izvedena od rodensa, participa prezenta rodere – „gristi“, „jesti“. Zečevi, zečevi i pike (red Lagomorpha) imaju kontinuirano rastuće sjekutiće, kao i glodari, i svojevremeno su bili uključeni u red. Međutim, oni imaju dodatni par sjekutića u gornjoj čeljusti i ova dva reda imaju prilično odvojenu evolucijsku historiju. Filogenija glodara svrstava ih u Glires, Euarchontoglires i Boreoeutheria. Slika ispod pokazuje unutrašnje i vanjske odnose Rodentia na osnovu pokušaja iz 2012. od strane Wua et al. da se uskladi molekularni sat s paleontološkim podacima [61]:
| Boreoeutheria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Red Rodentia može se podijeliti na podredove, infraredove, nadporodice i porodice. Postoji velika doza paralelizma i konvergencije među glodarima uzrokovana činjenicom da su imali tendenciju evolucije da popune uglavnom slične niše. Ova paralelna evolucija uključuje ne samo strukturu zuba, već i infraorbitalni dio lobanje (ispod očne duplje) i otežava klasifikaciju jer slične osobine možda nisu posljedica zajedničkog porijekla [62][63]. Brandt (1855) je prvi predložio podjelu glodara na tri podreda, Sciuromorpha, Hystricomorpha i Myomorpha, na osnovu razvoja određenih mišića u vilici i ovaj sistem je bio široko prihvaćen. Schlosser (1884) je izvršio sveobuhvatan pregled fosila glodara, uglavnom koristeći obrazne zube, i otkrio da se oni uklapaju u klasični sistem, ali Tullborg (1899) je predložio samo dva podreda, Sciurognathi i Hystricognathi. Oni su se zasnivali na stepenu savijanja donje vilice i trebali su biti dalje podijeljeni na Sciuromorpha, Myomorpha, Hystricomorpha i Bathyergomorpha. Matthew (1910) je stvorio filogenetsko stablo glodara Novog svijeta, ali nije uključio problematičnije vrste Starog svijeta. Dalji pokušaji klasifikacije nastavljeni su bez dogovora, pri čemu su neki autori usvojili klasični sistem od tri podreda, a drugi Tullborgova dva podreda.
Ova neslaganja ostaju neriješena, niti su molekularne studije u potpunosti riješile situaciju iako su potvrdile monofiliju grupe i da je poticala od zajedničkog paleocenskog pretka. Carleton i Musser (2005) u Sisarskim vrstama svijeta privremeno su usvojili sistem od pet podreda: Sciuromorpha, Castorimorpha, Myomorpha, Anomaluromorpha i Hystricomorpha. Od 2021. godine Američko društvo mamaloga prepoznaje 34 nedavne porodice koje sadrže više od 481 roda i 2277 vrsta [64][65].
Međuodnos sa ljudima
[uredi | uredi izvor]Zaštita
[uredi | uredi izvor]
Iako glodari nisu najozbiljnije ugroženi red sisara, postoji 168 vrsta u 126 rodova za koje se smatra da treba obratiti posebnu pažnju na njihovom očuvanju [66]. Budući da 76 posto rodova glodara sadrži samo jednu vrstu, velika filogenetska raznolikost mogla bi biti izgubljena s relativno malim brojem izumiranja. U nedostatku detaljnijeg znanja o ugroženim vrstama i tačne taksonomije, očuvanje se mora zasnivati uglavnom na višim taksonima (kao što su porodice, a ne vrste) i geografskim žarištima. Nekoliko vrsta pirinčanih pacova je izumrlo od XIX vijeka, vjerovatno zbog gubitka staništa i unošenja stranih vrsta [67]. U Kolumbiji je smeđi dlakavi patuljasti dikobraz zabilježen na samo dva planinska lokaliteta 1920-ih godina, dok je bodljasti štakor s crvenom dlakom poznat samo po svom tipskom lokalitetu na obali Kariba, pa se ove vrste smatraju ranjivim. IUCN Komisija za preživljavanje vrsta navodi "Možemo sa sigurnošću zaključiti da su mnogi južnoamerički glodari ozbiljno ugroženi, uglavnom zbog ekoloških poremećaja i intenzivnog lova" [68].
Tri vrste glodara [69], smeđi štakor, crni pacov i kućni miš proširile su se po svijetu tokom evropskih ekspedicija od XV do XIX vijeka. Pacifički polinezijski štakor (Rattus exulans) ozbiljno je oštetio ostrvske biote širom svijeta. Na primjer, kada je crni pacov stigao na ostrvo Lord Howe 1918. godine, preko 40 posto kopnenih vrsta ptica na ostrvu [70] je izumrlo u roku od deset godina. Slično uništenje viđeno je na ostrvu Midvej (1943.) i Velikom ostrvu Južni Kejp (1962.). Projekti očuvanja mogu uz pažljivo planiranje u potpunosti iskorijeniti ove invazivne glodare sa otoka korištenjem antikoagulansnog rodenticida kao što je brodifakum. Ovaj pristup je bio uspješan na ostrvu Lundy u Ujedinjenom Kraljevstvu, gdje je iskorjenjivanje otprilike 40.000 smeđih pacova dalo populaciji manks strižnjaka i atlantskog puffina priliku da se oporavi od skorog izumiranja.
Glodari su također podložni klimatskim promjenama, posebno vrste koje žive na niskim ostrvima. Bramble Cay melomys, koji je živio na najsjevernijoj tački kopna Australije, bio je prva vrsta sisara koja je proglašena izumrlom kao posljedica savremenih klimatskih promjena [71].
Iskorištavanje
[uredi | uredi izvor]
Čovječanstvo je dugo koristilo životinjske kože za odjeću, jer je koža izdržljiva, a krzno pruža dodatnu izolaciju. Domoroci Sjeverne Amerike mnogo su koristili dabrovu kožu, štaveći i šivajući je kako bi napravili ogrtače. Evropljani su cijenili njihov kvalitet i sjevernoamerička trgovina krznom se razvila i postala od primarne važnosti za prve naseljenike. U Evropi se pokazalo da je mekana donja vuna poznata kao "dabrova vuna" idealna za filcanje [72]. Kasnije je koypu postao jeftiniji izvor krzna za filcanje i bio je ekstenzivno uzgajan u Americi i Evropi; međutim, kako se moda se promijenila a novi materijali su postali dostupni, industrija životinjskog krzna se smanjila. Činčila ima mekanu i svilenkastu dlaku, a potražnja za njenim krznom bila je tolika da je skoro uništena u divljini prije nego što je uzgoj postao glavni izvor krzna.
Ishrana
[uredi | uredi izvor]Najmanje 89 vrsta glodara, uglavnom Hystricomorpha, kao što su zamorci, aguti i kapibare, jedu ljudi; 1985. postojala su najmanje 42 različita društva u kojima ljudi jedu pacove. Zamorci su prvi put uzgajani za hranu oko 2500 godina p.n.e. i do 1500. godine p.n.e. postali su glavni izvor mesa za Carstvo Inka. Rimljani su puhove uzgajali u posebnim posudama zvanim "gliraria", ili u velikim otvorenim ograđenim prostorima, gdje su ih tovili orasima, kestenovima i žirom. Puhovi su se također hvatali u divljini u jesen kada su bili najdeblji i pripremali pečeni. Istraživači su otkrili da su u Amazoniji, gdje su veliki sisari bili rijetki, pake i obični agouti činili oko 40 posto godišnje divljači koju su ulovili autohtoni ljudi, ali u šumskim područjima gdje su veći sisari bili prisutni u izobilju, ovi glodari su činili samo oko 3 posto ishrane [73].
Zamorci se koriste i u kuharstvu Kuska, Peru, u jelima kao što je cuy al horno, pečeni zamorac [74]. Tradicionalna andska peć, poznata kao qoncha ili fogón, napravljena je od blata i gline ojačane slamom i dlakom životinja kao što su zamorci. U Peruu je intenzivan uzgoj domaćih zamoraca. Ova životinja je odličan izvor hrane jer meso sadrži 19% proteina. U Sjedinjenim Državama ljudi jedu uglavnom vjeverice, ali i dikobraze i mrmote. Narod Navaho jeo je jelo pripremljeno od prerijskih pasa, dok su Paiute jeli gofove, vjeverice i pacove.
Testiranje na životinjama
[uredi | uredi izvor]Glodari se široko koriste kao uzorni organizmi u testiranju na životinjama [75]. Albino mutantni štakori su prvi put korišteni za istraživanja 1828. godine, a kasnije su postali prva životinja uzgajana za čisto naučne svrhe [76]. U današnje vrijeme kućni miš je najčešće korišteni laboratorijski glodar, a 1979. godine procijenjeno je da se godišnje koristi pedeset miliona primjeraka širom svijeta. Omiljeni su zbog svoje male veličine, plodnosti, kratkog perioda trudnoće i lakoće rukovanja i zato što su podložni mnogim stanjima i infekcijama koje pogađaju i ljude. Koriste se u istraživanju genetike, razvojne biologije, biologije ćelije, onkologije i imunologije. Zamorci su bili popularne laboratorijske životinje do kasnog XX vijeka; oko 2,5 miliona zamoraca godišnje je korišteno u Sjedinjenim Državama za istraživanje 1960-ih, ali se taj ukupan broj smanjio na oko 375,000 do sredine 1990-ih. U 2007. godini činile su oko 2% svih laboratorijskih životinja [77]. Zamorci su igrali glavnu ulogu u uspostavljanju teorije klica u kasnom XIX vijeku, kroz eksperimente Louisa Pasteura, Emilea Rouxa i Roberta Kocha. Nekoliko puta su lansirani u orbitalni svemirski let — prvo od strane SSSR-a na biosatelitu Sputnjik 9 dana 9. marta 1961. godine, uz uspješan povratak [78].
Glodari imaju osjetljive olfaktorne sposobnosti, koje su ljudi koristili za otkrivanje mirisa ili hemikalija od interesa. Gambijski štakor sa vrećicom je u stanju da detektuje bacile tuberkuloze sa osetljivošću do 86,6%, a otkrivanje odsustva bacila od preko 93%; ista vrsta je obučena za otkrivanje nagaznih mina. Pacovi su proučavani za moguću upotrebu u opasnim situacijama kao što su zone katastrofe. Mogu se obučiti da odgovaraju na komande, koje se mogu davati na daljinu, pa čak i uvjeriti da se upuste u jarko osvijetljena područja, što pacovi obično izbjegavaju [79].
Kao kućni ljubimci
[uredi | uredi izvor]Glodari uključujući zamorce, miševe, pacove, hrčke, gerbile, činčile, degue i vjeverice su pogodni kućni ljubimci koji mogu živjeti u malim prostorima. Većina se obično drži u kavezima odgovarajućih veličina i imaju različite zahtjeve za prostorom i društvenom interakcijom. Ako se njima rukuje od malih nogu, obično su poslušni i ne grizu. Zamorci imaju dug životni vijek i potreban im je veliki kavez. Štakorima je takođe potrebno dosta prostora i mogu postati veoma pitomi, mogu naučiti trikove i čini se da uživaju u ljudskom društvu. Miševi su kratkog vijeka, ali zauzimaju vrlo malo prostora. Hrčci su usamljeni, a imaju tendenciju da budu noću aktivni. Imaju zanimljivo ponašanje, ali ako se njima ne postupa na odgovarajući način, mogu biti odbrambenog ponašanja. Gerbili obično nisu agresivni, rijetko grizu i društvene su životinje koje uživaju u društvu ljudi i svoje vrste [80].
Kao štetočine i prenosioci bolesti
[uredi | uredi izvor]
Neke vrste glodara su ozbiljne poljoprivredne štetočine, jedu velike količine hrane koju proizvode ljudi [81]. Na primjer, 2003. godine procijenjeno je da je količina riže koju su miševi i pacovi pojeli u Aziji dovoljna da prehrani 200 miliona ljudi. Većinu štete širom svijeta uzrokuje relativno mali broj vrsta, uglavnom pacova i miševa. U Indoneziji i Tanzaniji glodari smanjuju prinose usjeva za oko petnaest posto, dok su u nekim slučajevima u Južnoj Americi gubici dosegli devedeset posto. Širom Afrike, glodari, uključujući Mastomys i Arvicanthis, oštećuju žitarice, kikiriki, povrće i kakao. U Aziji, pacovi, miševi i vrste kao što su Microtus brandti, Meriones unguiculatus i Eospalax baileyi oštećuju usjeve riže, sirka, gomolja, povrća i orašastih plodova. U Evropi, kao i pacovi i miševi, vrste Apodemus, Microtus i povremeno Arvicola terrestris nanose štetu voćnjacima, povrću i pašnjacima, te žitaricama. U Južnoj Americi, širi spektar vrsta glodara, kao što su Holochilus, Akodon, Calomys, Oligoryzomys, Phyllotis, Sigmodon i Zygodontomys, oštećuju mnoge usjeve uključujući šećernu trsku, voće, povrće i gomolje [82].
Glodari su također značajni prenosioci bolesti [83]. Crni štakor, sa buhama koje nosi, igra primarnu ulogu u širenju bakterije Yersinia pestis odgovorne za bubonsku kugu [84], i nosi organizme odgovorne za tifus, Weilovu bolest, toksoplazmozu i trihinelozu. Brojni glodari nose hantaviruse, uključujući viruse Puumala, Dobrava i Saaremaa, koji mogu zaraziti ljude. Glodavci također pomažu u prenošenju drugih bolesti uključujući babeziozu, kožnu lajšmaniozu, ljudsku granulocitnu anaplazmozu, lajmsku bolest, Omsku hemoragijsku groznicu, Powassan virus, rikecije, povratnu groznicu, pjegavu groznicu Rocky Mountaina i virus Zapadnog Nila.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ http://animaldiversity.ummz.umich.edu/site/accounts/information/Rodentia.html
- ^ Single, G.; Dickman, C. R.; MacDonald, D. W. (2001). "Rodents". In MacDonald, D. W. (ed.). The Encyclopedia of Mammals (2nd ed.). Oxford University Press. pp. 578–587. ISBN 978-0-7607-1969-5.
- ^ a b Waggoner, Ben (15 August 2000). "Introduction to the Rodentia". University of California Museum of Paleontology. Retrieved 4 July 2014.
- ^ Nowak, R. M. (1999). Walker's Mammals of the World. Johns Hopkins University Press. p. 1244. ISBN 978-0-8018-5789-8.
- ^ Single, G.; Dickman, C. R.; MacDonald, D. W. (2001). "Rodents". In MacDonald, D. W. (ed.). The Encyclopedia of Mammals (2nd ed.). Oxford University Press. pp. 578–587.
- ^ Cox, Philip G.; Rayfield, Emily J.; Fagan, Michael J.; Herrel, Anthony; Pataky, Todd C.; Jeffery, Nathan (27 April 2012). "Functional Evolution of the Feeding System in Rodents". PLOS ONE. 7 (4): e36299. Bibcode:2012PLoSO...736299C. doi:10.1371/journal.pone.0036299. ISSN 1932-6203. PMC 3338682. PMID 22558427.
- ^ Froberg-Fejko, Karen (1 October 2014). "Give a rat a bone: satisfying rodents' need to gnaw". Lab Animal. 43 (10): 378–379. doi:10.1038/laban.611. ISSN 1548-4475. S2CID 19686731.
- ^ Cox, Philip G.; Jeffery, Nathan (2011). "Reviewing the Morphology of the Jaw-Closing Musculature in Squirrels, Rats, and Guinea Pigs with Contrast-Enhanced MicroCT" (PDF). The Anatomical Record. 294 (6): 915–928. doi:10.1002/ar.21381. PMID 21538924. S2CID 17249666.
- ^ Mustapha, O. (2015). "Morphology of the Oral Cavity of the African Giant Rat". Bulgarian Journal of Veterinary Medicine. 18 (1): 19–30. doi:10.15547/bjvm.793.
- ^ horington, R. W Jr.; Darrow, K.; Anderson, C. G. (1998). "Wing tip anatomy and aerodynamics in flying squirrels" (PDF). Journal of Mammalogy. 79 (1): 245–250. doi:10.2307/1382860. JSTOR 1382860.
- ^ Petersen, Carl C.H. (8 July 2014). "Cortical Control of Whisker Movement". Annual Review of Neuroscience. Annual Reviews. 37 (1): 183–203. doi:10.1146/annurev-neuro-062012-170344. ISSN 0147-006X. PMID 24821429.
- ^ Horn, Charles C.; Kimball, Bruce A.; Wang, Hong; Kaus, James; Dienel, Samuel; Nagy, Allysa; Gathright, Gordon R.; Yates, Bill J.; Andrews, Paul L. R. (10 April 2013). Covasa, Mihai (ed.). "Why Can't Rodents Vomit? A Comparative Behavioral, Anatomical, and Physiological Study". PLoS ONE. 8 (4): e60537. Bibcode:2013PLoSO...860537H. doi:10.1371/journal.pone.0060537. ISSN 1932-6203. PMC 3622671. PMID 23593236.
- ^ Kapoor, Harit; Lohani, Kush Raj; Lee, Tommy H.; Agrawal, Devendra K.; Mittal, Sumeet K. (27 July 2015). "Animal Models of Barrett's Esophagus and Esophageal Adenocarcinoma-Past, Present, and Future". Clinical and Translational Science. 8 (6): 841–847. doi:10.1111/cts.12304. PMC 4703452. PMID 26211420.
- ^ Balaban, Carey D.; Yates, Bill J. (2017). "What is nausea? A historical analysis of changing views". Autonomic Neuroscience. 202: 5–17. doi:10.1016/j.autneu.2016.07.003. ISSN 1566-0702. PMC 5203950. PMID 27450627.
- ^ Schier, Lindsey A.; Spector, Alan C. (1 January 2019). "The Functional and Neurobiological Properties of Bad Taste". Physiological Reviews. 99 (1): 605–663. doi:10.1152/physrev.00044.2017. ISSN 0031-9333. PMC 6442928. PMID 30475657.
- ^ Schulte-Hostedde, A. I. (2008). "Chapter 10: Sexual Size Dimorphism in Rodents". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 117–119. ISBN 978-0-226-90538-9.
- ^ Helgen, Kristofer M. (2005). "The amphibious murines of New Guinea (Rodentia, Muridae): the generic status of Baiyankamys and description of a new species of Hydromys". Zootaxa. 913: 1–20. doi:10.11646/zootaxa.913.1.1. ISSN 1175-5326.
- ^ Parshad, V.R. (1999). "Rodent control in India" (PDF). Integrated Pest Management Reviews. 4 (2): 97–126. doi:10.1023/A:1009622109901. S2CID 36804001.
- ^ Krueger, Kirsten (1986). "Feeding relationships among bison, pronghorn, and prairie dogs: an experimental analysis". Ecology. 67 (3): 760–770. doi:10.2307/1937699. ISSN 0012-9658. JSTOR 1937699.
- ^ Pérez, Francisco; Castillo-Guevara, Citlalli; Galindo-Flores, Gema; Cuautle, Mariana; Estrada-Torres, Arturo (2012). "Effect of gut passage by two highland rodents on spore activity and mycorrhiza formation of two species of ectomycorrhizal fungi (Laccaria trichodermophora and Suillus tomentosus)". Botany. 90 (11): 1084–1092. doi:10.1139/b2012-086. ISSN 1916-2790.
- ^ Burchsted, D.; Daniels, M.; Thorson, R.; Vokoun, J. (2010). "The river discontinuum: applying beaver modifications to baseline conditions for restoration of forested headwaters". BioScience. 60 (11): 908–922. doi:10.1525/bio.2010.60.11.7. S2CID 10070184.
- ^ Wright, J. P.; Jones, C. G.; Flecker, A. S. (2002). "[www.caryinstitute.org/sites/default/files/public/reprints/Wright_et_al_2002_An_ecosystem_Oecologia_132_96-101.pdf An ecosystem engineer, the beaver, increases species richness at the landscape scale]". Oecologia. 132 (1): 96–101. Bibcode:2002Oecol.132...96W. doi:10.1007/s00442-002-0929-1. PMID 28547281. S2CID 5940275.
- ^ Kemp, P. S.; Worthington, T. A.; Langford, T. E. l.; Tree, A. R. J.; Gaywood, M. J. (2012). "Qualitative and quantitative effects of reintroduced beavers on stream fish". Fish and Fisheries. 13 (2): 158–181. doi:10.1111/j.1467-2979.2011.00421.x.
- ^ Hansson, Lennart (1971). "Habitat, food and population dynamics of the field vole Microtus agrestis (L.) in south Sweden". Viltrevy. 8: 268–278. ISSN 0505-611X. Archived from the original on 27 September 2013.
- ^ Attenborough, David (2002). The Life of Mammals. BBC Books. pp. 61–86. ISBN 978-0-563-53423-5.
- ^ Müller-Schwarze, Dietland; Sun, Lixing (2003). The Beaver: Natural History of a Wetlands Engineer. Cornell University Press. pp. 67–75. ISBN 978-0-8014-4098-4.
- ^ Landry, Stuart O. Jr. (1970). "The Rodentia as omnivores". The Quarterly Review of Biology. 45 (4): 351–372. doi:10.1086/406647. JSTOR 2821009. PMID 5500524. S2CID 30382320.
- ^ Jarvis, Jennifer (1981). "Eusociality in a mammal: Cooperative breeding in naked mole-rat colonies". Science. 212 (4494): 571–573. Bibcode:1981Sci...212..571J. doi:10.1126/science.7209555. JSTOR 1686202. PMID 7209555.
- ^ Hoogland, John L. (1995). The Black-Tailed Prairie Dog: Social Life of a Burrowing Mammal. University of Chicago Press. p. 1. ISBN 978-0-226-35118-6.
- ^ Baker, Bruce W.; Hill, Edward P. (2003). "Chapter 15: Beaver". In Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A. (eds.). Wild Mammals of North America: Biology, Management, and Conservation. JHU Press. pp. 288–310. ISBN 978-0-8018-7416-1.
- ^ Winslow, James T.; Hastings, Nick; Carter, C. Sue; Harbaugh, Carroll R.; Insel, Thomas R. (1993). "A role for central vasopressin in pair bonding in monogamous prairie voles" (PDF). Letters to Nature. 365 (6446): 545–548. Bibcode:1993Natur.365..545W. doi:10.1038/365545a0. PMID 8413608. S2CID 4333114.
- ^ Yensen, Eric; Sherman, Paul W. (2003). "Chapter 10: Ground Squirrels". In Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A. (eds.). Wild Mammals of North America: Biology, Management, and Conservation. JHU Press. pp. 211–225. ISBN 978-0-8018-7416-1.
- ^ Holmes, Warren G.; Mateo, Jill M. (2008). "Chapter 19: Kin Recognition in Rodents: Issues and Evidence". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 216–230. ISBN 978-0-226-90538-9.
- ^ Bjorkoyli, Tore; Rosell, Frank (2002). "A test of the dear enemy phenomenon in the Eurasian beaver". Animal Behaviour. 63 (6): 1073–1078. doi:10.1006/anbe.2002.3010. hdl:11250/2437993. S2CID 53160345.
- ^ Shelley, Erin L.; Blumstein, Daniel T. (2005). "The evolution of vocal alarm communication in rodents". Behavioral Ecology. 16 (1): 169–177. CiteSeerX 10.1.1.541.4408. doi:10.1093/beheco/arh148.
- ^ Zimmermann, Elke; Leliveld, Lisette; Schehka, Lisette (2013). "8: Toward the evolutionary roots of affective prosody in human acoustic communication: A comparative approach to mammalian voices". In Altenmüller, Eckart; Schmidt, Sabine; Zimmermann, Elke (eds.). The Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man. Oxford University Press. pp. 123–124. ISBN 978-0-19-164489-4.
- ^ Long, C. V. (2007). "Vocalisations of the degu (Octodon degus), a social caviomorph rodent". Bioacoustics. 16 (3): 223–244. doi:10.1080/09524622.2007.9753579. ISSN 0952-4622. S2CID 84569309.
- ^ Haverkamp, Silke; Waessle, Heinz; Duebel, Jens; Kuner, Thomas; Augustine, George J.; Feng, Guoping; Euler, Thomas (2005). "The primordial, blue-cone color system of the mouse retina". Journal of Neuroscience. 25 (22): 5438–5445. doi:10.1523/JNEUROSCI.1117-05.2005. PMC 6725002. PMID 15930394.
- ^ Desjardins, C.; Maruniak, J. A.; Bronson, F. H. (1973). "Social rank in house mice: Differentiation revealed by ultraviolet visualization of urinary marking patterns". Science. 182 (4115): 939–941. Bibcode:1973Sci...182..939D. doi:10.1126/science.182.4115.939. PMID 4745598. S2CID 44346136.
- ^ Viitala, J.; Korpimäki, E.; Palokangas, P.; Koivula, M. (1995). "Attraction of kestrels to vole scent marks visible in ultraviolet light". Nature. 373 (6513): 425–427. Bibcode:1995Natur.373..425V. doi:10.1038/373425a0. S2CID 4356193.
- ^ "Vibrational communication in mammals". Map of Life: Convergent evolution online. University of Cambridge. 4 August 2010.
- ^ Randall, J. A. (2001). "Evolution and function of drumming as communication in mammals". American Zoologist. 41 (5): 1143–1156. doi:10.1093/icb/41.5.1143.
- ^ Randall, Jan A.; Matocq, Marjorie D. (1997). "Why do kangaroo rats (Dipodomys spectabilis) footdrum at snakes?". Behavioral Ecology. 8 (4): 404–413. doi:10.1093/beheco/8.4.404.
- ^ Narins, P. M.; Reichman, O. J.; Jarvis, J. U. M.; Lewis, E. R. (1992). "Seismic signal transmission between burrows of the Cape mole-rat Georychus capensis". Journal of Comparative Physiology A. 170 (1): 13–22. doi:10.1007/BF00190397. PMID 1573567. S2CID 22600955.
- ^ Waterman, Jane (2008). "Chapter 3: Male Mating Strategies in Rodents". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 28–39. ISBN 978-0-226-90538-9.
- ^ McGuire, Betty; Bernis, William E. (2008). "Chapter 20: Parental Care". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 231–235. ISBN 978-0-226-90538-9.
- ^ Holmes, Warren G.; Mateo, Jill M. (2008). "Chapter 19: Kin Recognition in Rodents: Issues and Evidence". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 226–227. ISBN 978-0-226-90538-9.
- ^ Ebensperger, Luis A.; Blumsperger, Daniel T. (2008). "Chapter 23: Nonparental Infanticide". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 274–278. ISBN 978-0-226-90538-9.
- ^ Hackländera, Klaus; Möstlb, Erich; Arnold, Walter (2003). "Reproductive suppression in female Alpine marmots, Marmota marmota". Animal Behaviour. 65 (6): 1133–1140. doi:10.1006/anbe.2003.2159. S2CID 53218701.
- ^ McGuire, Betty; Bernis, William E. (2008). "Chapter 20: Parental Care". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 231–235. ISBN 978-0-226-90538-9.
- ^ Jacobs, Lucia F. (1992). "Memory for cache locations in Merriam's kangaroo rats" (PDF). Animal Behaviour. 43 (4): 585–593. doi:10.1016/S0003-3472(05)81018-8. S2CID 14173113.
- ^ Harding, E. J.; Paul, E. S.; Mendl, M. (2004). "Animal behaviour: Cognitive bias and affective state". Nature. 427 (6972): 312. Bibcode:2004Natur.427..312H. doi:10.1038/427312a. PMID 14737158. S2CID 4411418.
- ^ Foote, Allison L.; Crystal, J. D. (2007). "Metacognition in the rat". Current Biology. 17 (6): 551–555. doi:10.1016/j.cub.2007.01.061. PMC 1861845. PMID 17346969.
- ^ Jozefowiez, J.; Staddon, J. E. R.; Cerutti, D. T. (2009). "Metacognition in animals: how do we know that they know?". Comparative Cognition & Behavior Reviews. 4: 29–39. doi:10.3819/ccbr.2009.40003.
- ^ Kay, Emily H.; Hoekstra, Hopi E. (2008). "Rodents". Current Biology. 18 (10): R406–R410. doi:10.1016/j.cub.2008.03.019. PMID 18492466.
- ^ Horner, D. S.; Lefkimmiatis, K.; Reyes, A.; Gissi, C.; Saccone, C.; Pesole, G. (2007). "Phylogenetic analyses of complete mitochondrial genome sequences suggest a basal divergence of the enigmatic rodent Anomalurus". BMC Evolutionary Biology. 7 (1): 16. doi:10.1186/1471-2148-7-16. PMC 1802082. PMID 17288612.
- ^ Schenk, John J.; Rowe, Kevin C.; Steppan, Scott J. (2013). "Ecological opportunity and incumbency in the diversification of repeated continental colonizations by muroid rodents". Systematic Biology. 62 (6): 837–864. doi:10.1093/sysbio/syt050. PMID 23925508.
- ^ Marivaux, Laurent; Essid, El Mabrouk; Marzougui, Wissem; Ammar, Hayet Khayati; Adnet, Sylvain; Marandat, Bernard; Merzeraud, Gilles; Tabuce, Rodolphe; Vianey-Liaud, Monique (2014). "A new and primitive species of Protophiomys (Rodentia, Hystricognathi) from the late middle Eocene of Djebel el Kébar, Central Tunisia". Palaeovertebrata. 38 (1): 1–17. doi:10.18563/pv.38.1.e2. S2CID 55599571.
- ^ Rowe, K. C.; Reno, M. L.; Richmond, D. M.; Adkins, R. M.; Steppan, S. J. (2008). "Pliocene colonization and adaptive radiations in Australia and New Guinea (Sahul): multilocus systematics of the old endemic rodents (Muroidea: Murinae)". Molecular Phylogenetics and Evolution. 47 (1): 84–101. doi:10.1016/j.ympev.2008.01.001. PMID 18313945.
- ^ Marshall, L. G.; Butler, R. F.; Drake, R. E.; Curtis, G. H.; Tedford, R. H. (1979). "Calibration of the Great American Interchange". Science. 204 (4390): 272–279. Bibcode:1979Sci...204..272M. doi:10.1126/science.204.4390.272. PMID 17800342. S2CID 8625188.
- ^ Wu, Shaoyuan; Wu, Wenyu; Zhang, Fuchun; Ye, Jie; Ni, Xijun; Sun, Jimin; Edwards, Scott V.; Meng, Jin; Organ, Chris L. (2012). "Molecular and paleontological evidence for a post-Cretaceous origin of rodents". PLOS ONE. 7 (10): e46445. Bibcode:2012PLoSO...746445W. doi:10.1371/journal.pone.0046445. PMC 3465340. PMID 23071573.
- ^ Wood, Albert E. (1955). "A Revised Classification of the Rodents". Journal of Mammalogy. 36 (2): 165–187. doi:10.2307/1375874. JSTOR 1375874.
- ^ Wood, Albert E. (1958). "Are there rodent suborders?". Systematic Biology. 7 (4): 169–173. doi:10.2307/2411716. JSTOR 2411716.
- ^ Carleton, M. D.; Musser, G. G. (2005). "Order Rodentia". In Wilson, Don E.; Reeder, DeeAnn M. (eds.). Mammal Species of the World – a taxonomic and geographic reference. Vol. 12. JHU Press. pp. 745–752. ISBN 978-0-8018-8221-0.
- ^ Honeycutt, Rodney L. (2009). "Rodents (Rodentia)" (PDF). In Hedges, S.B.; Kumar, S. (eds.). The Timetree of Life. Oxford University Press.
- ^ mori, G.; Gippoliti, S. (2003). "A higher-taxon approach to rodent conservation priorities for the 21st century". Animal Biodiversity and Conservation. 26 (2): 1–18.
- ^ Morgan, G. S. (1993). "Quaternary land vertebrates of Jamaica". Biostratigraphy of Jamaica. Geological Society of America Memoirs. Vol. 182. pp. 417–442. doi:10.1130/mem182-p417. ISBN 978-0-8137-1182-9.
- ^ Gudynas, Eduardo (1989). Lidicker, William Zander (ed.). Rodents: A World Survey of Species of Conservation Concern: Based on the Proceedings of a Workshop of the IUCN/SSC Rodent Specialist Group, Held at the Fourth International Theriological Congress, August 17, 1985, Edmonton, Alberta, Canada. IUCN. p. 23.
- ^ Buckle, A. P.; Fenn, M. G. P. (1992). "Rodent Control in the Conservation of Endangered Species". Proceedings of the 15th Vertebrate Pest Conference: Paper 12. 3–5 March 1992
- ^ Hindwood, K.A. (1940). "Birds of Lord Howe Island". Emu. 40: 1–86. doi:10.1071/mu940001.
- ^ Slezak, Michael (14 June 2016). "Revealed: first mammal species wiped out by human-induced climate change". The Guardian. London.
- ^ Innis, Harold A. (1999). The Fur Trade in Canada: An Introduction to Canadian Economic History. University of Toronto Press. pp. 9–12. ISBN 978-0-8020-8196-4.
- ^ Fiedler, Lynwood A. (1990). "Rodents as a Food Source". Proceedings of the Fourteenth Vertebrate Pest Conference 1990: 149–155.
- ^ Knowlton, David (13 July 2011). "Guinea Pig, Pet or Festive Meal". Cuzco Eats. Archived from the original on 14 July 2014. Retrieved 5 July 2014. Morveli, Walter Coraza; Knowlton, David (5 March 2012). "Traditional Mud
- ^ Wolff, Jerry O.; Sherman, Paul W. (2008). Rodent Societies: An Ecological and Evolutionary Perspective. University of Chicago Press. pp. 3–8. ISBN 978-0-226-90538-9.
- ^ Krinke, George J.; Bullock, Gillian R.; Bunton, Tracie (2000). "History, strains and models". The Laboratory Rat (Handbook of Experimental Animals). Academic Press. pp. 3–16. ISBN 978-0-12-426400-7.
- ^ Gad, Shayne C. (2007). Animal Models in Toxicology (2nd ed.). Taylor & Francis. pp. 334–402. ISBN 978-0-8247-5407-5.
- ^ Gray, Tara (1998). "A Brief History of Animals in Space". National Aeronautics and Space Administration.
- ^ Xu, S.; Talwar, S. K.; Hawley, E. S.; Li, L.; Chapin, J. K. (2004). "A multi-channel telemetry system for brain microstimulation in freely roaming animals". Journal of Neuroscience Methods. 133 (1–2): 57–63.
- ^ Broekel, Ray (1983). Gerbil Pets and Other Small Rodents. Childrens Press. pp. 5–20. ISBN 978-0-516-01679-5.
- ^ Meerburg, B. G.; Singleton, G. R; Leirs, H. (2009). "The Year of the Rat ends: time to fight hunger!". Pest Management Science. 65 (4): 351–2. doi:10.1002/ps.1718. PMID 19206089.
- ^ Stenseth, Nils Chr; Leirs, Herwig; Skonhoft, Anders; Davis, Stephen A.; Pech, Roger P.; Andreassen, Harry P.; Singleton, Grant R.; Lima, Mauricio; Machang'u, Robert S.; Makundi, Rhodes H.; Zhang, Zhibin; Brown, Peter R.; Shi, Dazhao; Wan, Xinrong (2003). "Mice, rats, and people: The bio-economics of agricultural rodent pests". Frontiers in Ecology and the Environment. 1 (77): 367–375. doi:10.2307/3868189. JSTOR 3868189.
- ^ Meerburg, B. G.; Singleton, G. R.; Kijlstra, A. (2009). "Rodent-borne diseases and their risks for public health". Critical Reviews in Microbiology. 35 (3): 221–70. doi:10.1080/10408410902989837. PMID 19548807. S2CID 205694138.
- ^ McCormick, M. (2003). "Rats, communications, and plague: Toward an ecological history" (PDF). Journal of Interdisciplinary History. 34 (1): 1–25. doi:10.1162/002219503322645439. S2CID 128567627.