Morfologija insekata

2. Ocele (donje)
3. Ocele (gornje)
4. Složeno oko
5. Mozak (moždana ganglija)
6. Protoraks
7. Leđni krvni sud
8. Trahejske cijevi (deblo sa spirakulumom)
9. Mezotoraks
10. Metatoraks
11. Prednje krilo
12. Stražnje krilo
13. Srednje crijevo (želudac)
14. Leđna cijev (srce)
15. Jajnik
16. Stražnje crijevo (tanko crijevo, rektum i anus)
17. Anus
18. Jajovod
19. Nervna vrpca (trbušna ganglija)
20. Malpighijeve cijevi
21. Stopalni jastučići
22. Kandže
23. Stopalo
24. Tibija
25. Femur
26. Trohanter
27. Prednje crijevo (voljka, želudac)
28. Grudna ganglija
29. Koksa
30. Pljuvačna žlijezda
31. Podjednjačka ganglija
32. Usni aparat
Morfologija insekata proučava i opisuje vanjska fizička svojstva insekata. Terminologija koja se koristi za opisivanje insekata slična je onoj koja se koristi za druge artropode zbog njihove zajedničke evolucijske povijesti. Tri fizička obilježja odvajaju insekte od ostalih člankonožaca: imaju tijelo podijeljeno u tri regije (glava, grudni koš i trbuh), imaju tri para nogu i dijelove usta koji se nalaze izvan glavene kapsule. Upravo ih taj položaj usnih dijelova dijeli od najbližeg srodnika, neinsekata šesteronožaca, uključujući i Protura, Diplura i Collembola.
Postoje ogromne razlike u strukturi tijela među vrstama insekata. Pojedini mogu biti veliki u rasponu od 0,3 mm (vilinska muha) do 30 cm (moljac velika sova);[1] nemaju očiju ili ih nema puno; imaju dobro razvijena krila ili ih nemaju; noge su prilagođene trčanju, skakanju, plivanju ili čak kopanju. Te preinake omogućuju insektima da zauzimaju gotovo svaku ekološku nišu na planeti, osim dubokog okeana. Ovaj članak opisuje osnovno tijelo insekata i neke od glavnih varijacija njegovih različitih dijelova. U tom procesu definira mnoge tehničke izraze koji se koriste za opisivanje tijela insekata.
Pregled anatomije
[uredi | uredi izvor]Insekti, kao i svi člankonošci, nemaju unutrašnji kostur (endoskelet); umjesto toga imaju egzoskelet, tvrdi vanjski sloj izrađen uglavnom od hitina, koji štiti i podupire tijelo. Tijelo insekata podijeljeno je na tri dijela: glava, toraks i trbuh (zadak).[2] Glava je specijalizirana za senzorne unose i unos hrane; grudni koš, koji je sidrište nogu i krila (ako ih ima), specijaliziran je za kretanje, a trbuh za probavu, disanje, izlučivanje i razmnožavanje.[1] Iako je opća funkcija triju tjelesnih regija jednaka za sve vrste insekata, postoje velike razlike u osnovnoj strukturi, u krilima, nogama, antenama i dijelovima usta koji se vrlo razlikuju od skupine do skupine.[3]
Vanjština
[uredi | uredi izvor]Egzoskelet
[uredi | uredi izvor]Vanjski skelet insekata, kutikula, sastoji se od dva sloja; epikutikula, koja je tanak, voštani, vodootporni vanjski sloj i ne sadrži hitin, a sloj ispod njega naziva se prokutikula. Ona je hitinska i mnogo je deblja od epikutikule i ima dva sloja, vanjski je egzokutikula, dok je unutrašnji endokutikula. Čvrsta i fleksibilna endokutikula građena je od brojnih slojeva vlaknastog hitina i proteina, međusobno se križajući u sendviču, dok je egzokutikula kruta i sklerotizirana.[1] Egzokutikula je znatno reducirana kod mnogih insekata mehkog tijela, posebno u fazama larve (npr. gusjenica. Hemijski, hitin je dugolančani polimer N-acetilglukozamina, derivata glukoze. U svom nepromijenjenom obliku hitin je proziran, savitljiv, elastičan i prilično žilav. Međutim, u artropodima, on se često modificira, ugrađujući se u očvrsnuti protein acetatni matriks, koji čini velik dio egzoskeleta. U svom je čistom obliku kožnat, ali kada je obložen kalcij-karbonatom, postaje puno tvrđi.[4] Razlika između nemodificiranog i modificiranog oblika može se vidjeti usporedbom tjelesnog zida gusjenica (nemodificirane) s tvrdokrilcima (modificirane).
Iz samog embrionskog stadija, sloj stubastih ili kuboidnih epitelnih ćelija stvara vanjsku kutikulu i unutrašnju bazalnu membranu. Većina materijala insekata nalazi se u endokutikuli. Kutikula pruža mišićnu potporu i djeluje kao zaštitni štit dok se insdekt razvija. Međutim, budući da ne može rasti, vanjski sklerotizirani dio kutikule povremeno se odbacuje u procesu koji se naziva "presvlačenje" (mitarenje). Kako se vrijeme presvlačenja približava, većina egzokutikulnog materijala ponovno se upija. U mitrenju se najprije stara kutikula odvaja od epiderme (apoliza). Između stare kutikule i epiderme oslobađa se enzimska tekućina za odvajanje, koja razdvaja egzokutikulu probavljajući endokutikulu i odvajajući njen materijal za novu kutikulu. Kad se nova kožica dovoljno formira, epikutikula i reducirana egzokutikula izbacuju se u procesu zvanom ekdiza.[5]:16–20
Četiri glavna područja tjelesnih segmenata insekata su: tergum ili leđno, sternum ili trbušno i dvije pleure ili bočna. Stvrdnute ploče u egzoskeletu nazivaju se skleritima, koji su poddijelovi glavnih regija – tergita, sternita i pleurita, za odgovarajuće regije tergum, sternum i pleuron.[6]
Glava
[uredi | uredi izvor]
a: antenna,
b: ocele,
c: verteks,
d: složeno oko,
e: okciput,
f: gena,
g: pleurostoma,
h: mandibla ,
i: usnena palpa,
j: gornjevilične palpe ,
k: maksila,
l: labrum,
m: klipeus,
n: frons


Glava većine insekata zatvorena je u tvrdu, jako sklerotiziranu, egzoskeletnu glavenu kapsulu. Glavna je iznimka kod vrsta čija larva nije u potpunosti sklerotizirana, što se uglavnom javlja kod nekih holometabolnih. Međutim, čak i većina nesklerotiziranih ili slabo sklerotiziranih larvi često imaju dobro sklerotizirane kapsule glave, naprimjer larve Coleoptera i Hymenoptera. Larve Cyclorrhapha, međutim, nemaju gotovo nikakvu glavenu kapsulu.
Kapsula nosi većinu glavnih čulnih organa, uključujući antene, ocele i složene oči. Na njoj je također i usni aparat. U odraslog insekta glavena kapsula očito je nesegmentirana, premda embriološke studije pokazuju da se sastoji od šest segmenata koji nose uparene dodatke glave, uključujući dijelove usta, svaki par na određenom segmentu.[7] Svaki takav par zauzima jedan segment, iako svi segmenti modernih insekata ne nose vidljive dodatke.
Od svih redova insekata, Orthoptera najprikladnije prikazuje najveću raznolikost obilježja pronađenih na glavama insekata, uključujući suture i sklerite. Ovdje se verteks ili vrh (leđna regija) kod insekata s hipognatnim i opistognatnim glavama nalazi između složenih očiju. Kod insekata prognatizmom vrh se ne nalazi između složenih očiju, već tamo gdje se obično nalaze ocele. To je zato što je primarna os glave zakrenuta za 90°, kako bi postala paralelna s primarnom tjelesnom osovinom. Kod nekih je vrsta, ovo područje modificirano je i poprima drugo ime.[8]
Ekdizijska sutura (šav) sastoji se od kruničnih, frontalnih i epikranijalnih šavova, plus ekdizijalnih linija i linija cijepanja, koje se razlikuju među različitim vrstama insekata. Ekdizijalni šav uzdužno je postavljen na vrh i razdvaja epikranijske polovine glave s lijeve i desne strane. Ovisno o insektu, šav može biti u različitim oblicima: Y, U ili V. Te razilazeće linije koje čine ekdizijski šav nazivaju se frontalni ili 'frontogene suture. Nemaju sve vrste insekata frontalne suture, ali kod onih koji ih imaju, one se razdvajaju tokom ekdize, što pomaže da se otvori tjelesni omotač (integument) nove jedinke.
Frons je onaj dio glavene kapsule glave koji je ventralno ili ispred tjemena (verteksa). Veličina mu varira u odnosu na insekte, a kod mnogih vrsta definicija granica je proizvoljna, čak i kod nekih taksona koji imaju dobro definirane glavene kapsule. Međutim, kod većine vrsta fronsovi su sprijeda omeđeni frontoklipnim ili epistomnim sulkusom iznad klipeusa. Bočno je ograničen frontogenim sulkusom, ako je prisutan, i granicom s verteksom, ekdizijskom linijom cijepanja, ako je vidljiva. Ako postoji srednja ocela, ona je uglavnom na prednjim dijelovima, iako se kod nekih insekata, poput mnogih Hymenoptera, sva tri ocele pojavljuju na verteksu. Formalnija definicija je da je to sklerit iz kojeg proizlaze mišići dilatatori ždrijela, ali u mnogim kontekstima ni to nije primjenjivo.[7] IU anatomiji nekih taksona, poput mnogih Cicadomorpha, prednji dio glave prilično je jasno razlučen i ima tendenciju da bude širok i podokomit; za to se srednje područje obično uzima kao frons.[9]

Klipeus je sklerit između lica i labruma, koji je frontoklipeusnim šavom kod primitivnih insekata dorzalno odvojen od fronsa. Klipeogeni šav bočno razgraničava klipeus, pri čemu je klipeus ventralno odvojen od labruma klipeolabrumskim šavom. Klipeus se razlikuje u obliku i veličini, kao, što su vrste Lepidoptera s velikim klipeusom s izduženim dijelovima usta. Obraz ili gena formira sklerotizirano područje sa svake strane glave, ispod složenih očiju koje se protežu do šava ploča. Kao i mnogi drugi dijelovi koji čine glavu insekta, i gena se razlikuje među vrstama, čije je granice teško ustanoviti. Naprimjer, u Odonata nalazi se između složenih očiju, klipeusa i usnog aparata. Postgena je područje neposredno iza, ili straga ili niže na geni krilatih insekata ( Pterygota), a čini bočni i trbušni dio potiljačnog luka. Taj luk je uska traka stražnjeg rub glavene capsule, koji se dorzalno nadvija nad foramenom. Podgensko područje je obično usko, smješteno iznad dijelova usta; ono također uključuje hipostom i pleurostom.[8] Verteks se proteže sprijeda iznad dna antena kao istaknuti, šiljasti, udubljeni rostrum. Stražnji zid glavene kapsule prodire kroz veliki otvor, foramen. Kroz njega prolaze sistemi organa, poput nervna vrpca, jednjak, pljuvačni kanal i muskulatura, povezujući glavu sa toraks.[10]
Na zadnjoj površini glave su okciput, postgena, [[potiljačni otvor], stražnja tentorna jama, gula, postgeni most, [[hipostomska sutura] ] i most i donja vilica, labijum i maksila . Okcipitalna sutura dobro je utemeljena kod vrsta reda Orthoptera, ali ne toliko kod drugih redova. Tamo gdje je pronađena, potiljačni šav je lučni utor u obliku potkove na stražnjoj strani glave, koji završava na stražnjem dijelu svake donje vilice. Postpotiljačni šav je orijentir na stražnjoj površini glave i obično je u blizini potiljačnih otvora. Kod krilatih insekata, postokciput čini krajnji stražnji dio, često u obliku U, koji čini rub glave, protežuči se do postokcipitalnog šava. Kod njih, poput onih kod reda Orthoptera, potiljačni foramen i usta nisu odvojeni. Tri vrste okcipitalnih zatvarača ili tačaka ispod okcipitalnog foramena koje razdvajaju dvije donje polovine postgene su: hipostomski most, postgenalni most i gula. Hipostomalni most se obično nalazi kod insekata hipognetne orijentacije. Postgenalni most nalazimo kod odraslih vrsta viših Diptera i akuleatnih Hymenoptera, dok je gula na nekim Coleoptera, Neuroptera i Isoptera , koji tipski imaju usne dijelove prognatno (istureno) orijentirane.[8]
Složene oči i ocele
[uredi | uredi izvor]Većina insekata ima jedan par velikih, istaknutih složenih očiju, sastavljenih od jedinica zvanih ommatidije, moguće do 30.000 u jednom složenom oku, naprimjer, velikih vretenca. Ovaj tip oka postiže manju rezoluciju od očiju kod kičmenjaka, ali ima akutnu percepciju kretanja i obično je osjetljiv na UV-zrake i zelenu boju, a može imati dodatne vrhove osjetljivosti u drugim regijama vidnog spektra. Često imaju sposobnost detekcije E-vektora polarizovane svjetlosti polarizirane svjetlosti..[11] Mogu imati i dvije ili tri dodatne ocele, koje pomažu u otkrivanju slabe svjetlosti ili malih promjena u njenom intenzitetu. Opažena slika kombinacija je ulaza iz brojnih omatidija, smještenih na konveksnoj površini, usmjeravajući je tako u nešto drugačijim smjerovima. U poređenju sa jednostavnim očima, složene oči imaju vrlo velike uglove gledanja i bolju oštrinu od leđnih ocela insekata, ali neke matične materije (= oči larve), naprimjer one larve porodice Tenthredinidae s oštrinom od 4 stepena i vrlo visoka polarizacijska osjetljivost, odgovara performansama složenih očiju.[12] [13]


Budući da su pojedinačna sočiva tako mala, efekti difrakcije nameću ograničenje moguće rezolucije koja se može dobiti (pod pretpostavkom da ne funkcioniraju kao fazni niz. Ovome se može suprotstaviti samo povećanje veličine i broja sočiva. Da bi se vidjela rezolucija uporediva s ljudskim jednostavnim očima, ljudima bi trebale takve složene oči od kojih bi svako dosezalo veličinu glave. Složene oči dijele se u dvije skupine: apozicijske oči koje čine višestruko obrnute slike i superpozicijske oči koje vide jedostavu uspravnu sliku.[14][15] Compound eyes grow at their margins by the addition of new ommatidia.[16]
Antene
[uredi | uredi izvor]
Antene , koji se ponekad nazivaju i "osjetnici", fleksibilni su dodaci na glavi insekta, koji se koriste za otkrivanje okoliša. Insekti antenama "mogu" osjetiti pomoću finih dlačica (seta) koje ih prekrivaju.[17] Međutim, dodir nije jedini podražaj koji antene mogu otkriti; brojne sitne čulne strukture na antenama omogućavaju insektima da osjete mirise, temperaturu, vlažnost, pritisak, pa čak i potencijalno osjećaju sebe u prostoru.[17]:8–11[18][19] Some insects, including bees and some groups of flies can also detect sound with their antennae.[20]
Broj segmenata u anteni znatno varira među insektima, s tim što više muhe imaju samo 3-6 segmenata,[21] dok odrasli žohari mogu imati preko 140.[22] Opći oblik antena također je prilično varijabilan, ali prvi segment (onaj pričvršćen za glavu) uvijek se naziva stablo, a drugi segment pedicel. Preostali antenski segmenti ili flagelomere nazivaju se bičevima.[17]:8–11
Opći tipovi antena insekta prikazani su u nastavku:
 Aristatne |
 Kapitatne |
 Klavatne |
 Filiformne |
 Flabelatne |
 Genikulatne |
 Setatne |
 Lamelatne |
 Moniliformne |
 Pektinatne |
 Plumozne |
 Seratne |
 Stilatne |
Usni aparat
[uredi | uredi izvor]Usni aparat insekata sastoji se od maksile, labija (usana), a kod nekih vrsta i donje vilice (mandibule).[8][23] Labrum je jednostavan, srasli sklerit, koji se često naziva gornja usna, pruža se uzdužno i spojen je na klipeus. Mandibule (donje vilice) su visoko sklerotizirani par struktura koje se smještene pod pravim uglom u odnosu na tijelo i koriste se za grizenje, žvakanje i rezanje hrane. Maksile su uparene strukture koje se također mogu pružati pod pravim uglom u odnosu na tijelo i imaju segmentirane palpe. Labijum (donja usna) je srasla struktura koja se proteže uzdužno i ima par segmentiranih palpi.[24]
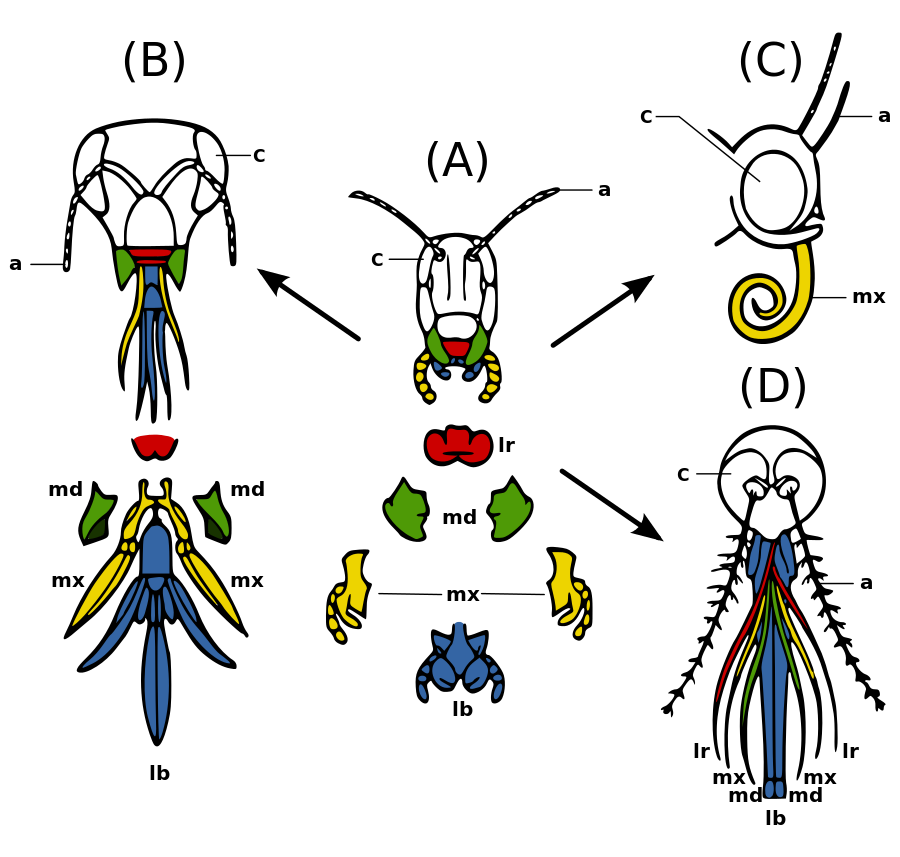
(A) primitivni aparat za žvakakanje kod skakavca, u centru
(B) tip za srkanje medonosne pčele,
(C) tip sifona leptira i
(D) za sisanje ženki komaraca.
a – antene,
c – složeno oko,
lb – labija;
lr – labrum;
md – mandibule,
mx – maksile
hp – hipofarinks
Dijelovi usnnog aparata, zajedno s ostatkom glave, mogu se zglobiti u najmanje tri različita položaja: prognatni, opistognatni i hipognatni. U vrsta s prognatnim zglobljavanjem, glava je postavljena okomito poravnato s tijelom, kao što su vrste porodice Formicidae (mrava); dok je kod hipognatnog tipa glava poravnata vodoravno uz tijelo. Opistognatna glava postavljena je dijagonalno, poput vrsta reda Blattodea i nekih Coleoptera.[25] Usni aparat se uveliko razlikuju među insektima različitih redova, ali dvije glavne funkcionalne skupine su mandibulatni i haustelatni. Hauselatni usni aparati koriste se za isisavanje tekućina i mogu se dalje klasificirati prema prisustvu stileta, što uključuje probadanje-sisanje, srkanje i sifoniranje. Stilete su ispupčenja nalik iglama koja se koriste za prodiranje u biljna i životinjska tkiva. One i cijev za hranjenje čine modificirane donje vilice, maksilu i hipofarinks.
- Mandibulatni dijelovi usnog aparata, među najčešćim su u insekata, a koriste se za grizenje i mljevenje čvrste hrane.
- Usni aparat za probadanje-sisanje ima stilete i koristi se za prodiranje u čvrsto tkivo, a zatim usisavanje tečne hrane.
- Spužvasti aparati koriste se za upijanje i sisanje tekućina, a nedostaju im stilete (npr. većina Diptera.
- Sifonski usni aparati nemaju stilete i koriste se za sisanje tečnosti, a često se nalaze među vrstama Lepidoptera.
Mandibulatni usni dijelovi nalaze se u vrstama Odonata, odraslih Neuroptera, Coleoptera, Hymenoptera, Blattodea, Orthoptera i Lepidoptera . Međutim, većina odraslih Lepidoptera ima usisne dijelove za usta, dok njihove larve (obično zvane gusjenice] imaju donje vilice (mandibule).
Mandibulatni
[uredi | uredi izvor]Labrum je široki režanj koji tvori krov predusne šupljine, obješen o klipeus ispred usta i čineći gornju usnu.[1] Na svojoj unutrašnjoj strani je opnast i može se pretvoriti u srednji režanj, epifarinks, koji nosi neke senzile. Labrum se podiže od donje vilice pomoću dva mišića koji polaze u glavi sa hvatištem medijalno na prednjem rubu labruma. Djelimično ga zatvaraju dva mandibulska mišića, sa polazištem u glavi i hvatištem na straznjim bočnim rubovima dva mala sklerita, tormama. Barem kod nekih insekata, hvatište je na rezlinskoj sponi na kutikuli, na spoju labruma sa klipeusom.[26] Donedavno se labrum općenito smatrao povezanim s prvim segmentom glave. Međutim, nedavna ispitivanja embriologije, ekspresije gena i opskrbe labruma živcem pokazuju da ga inervira tritocerebrum moždane ganglije, koja je srasla ganglija trećeg segmenta glave. To je nastalo spajanjem dijelova para dodataka predaka koji se nalaze na trećem segmentu glave, pokazujući njihov odnos. Njegova trbušna ili unutrašnja površina obično je opnasta i oblikuje epifarinks poput režnja koji nosi mehanosenzile i hemosenzile.[27][28]
Insektikoji žvaču imaju dvije donje vilice, po jednu sa svake strane glave. Mandibule su smještene između labruma i maksila. Mandibule režu i drobe hranu i mogu se koristiti za odbranu; uglavnom imaju apikalnu reznu oštricu i hranu melje više baznog viličnog područja. Mogu biti izuzetno tvrdi (oko 30 kg/mm2); prema tome, mnogi termiti i tvrdokrilci nemaju fizičkih poteškoća u probijanju kroz folije izrađene od uobičajenih metala, kao što su bakar, olovo, kalaj i cink.[1] Rezne ivice su obično ojačane dodatkom cinka, mangana ili rijetko željeza u količinama do oko 4% suhe mase. Oni su obično najveći dijelovi usnog aparata za žvakanje insekata , koji se koristi za žvakanje (rezanje, kidanje, drobljenje, mrvljenje) prehrambenih proizvoda. Otvaraju se prema van (sa strane glave) i međusobno se spajaju. U mesožderskih insekata koji žvaču, mandibule se mogu modificirati da budu sličnije noževima, dok su kod biljojednih uobičajenije široke i ravne na suprotnim obrazima (npr. gusjenica. Kod mužjaka jelenka (red Coleptera) donje vilice su modificirane do te mjere da ne služe nikakvoj hranidbenoj funkciji, već se umjesto toga koriste za odbranu mjesta parenja od drugih mužjaka. Kod mrava, donje vilice također imaju odbrambenu funkciju (posebno u vojničkim kastama). U bikovskih mrava mandibule su izdužene i nazubljene, a koriste se kao lovni (i odbrambeni) dodaci.
Smještene ispod donjih vilica, uparena maksile manipuliraju hranom tokom žvakanja. Maksile mogu imati dlake i "zube" duž unutrašnjih rubova. Na vanjskoj margini, galea je čašasta ili lopatasta struktura koja se nalazi preko vanjskog ruba labijuma. Oni takođe imaju palpe , koje se koriste za otkrivanje karakteristika potencijalne hrane. Maksile zauzimaju bočni položaj, po jedna sa svake strane glave iza donje vilice. Proksimalni dio maksile sastoji se od baznog kardoa koji ima jedan zglob s glavom i ravne ploče vilica, zakačene za kardo. I kardo i stipule su membranom labavo povezani s glavom, tako da se mogu kretati. Distalno na zidovima nalaze se dva režnja, unutrašnji lacinea i vanjski galea, od kojih jedan ili oba mogu biti odsutni. Na bočnim stranama je bočno spojena dlaka nalik nogama, sastavljena od niza segmenata. Kod Orthoptera ih je pet. Prednji i stražnji rotatorni mišići imaju hvatište na kardou, a trbušni aduktorski mišići koji polaze na tentoriju i aju hvatište na kardou i na zidovima. U stipesima polaze mišići fleksori lacinee i galeje, a drugi lacinealni fleksor ima polazište u lobanji, ali ni lacinea ni galea nemaju ekstenzorski mišić. Palpe imaju mišiće podizače i depresore, koji se pojavljuju u zidovima, a svaki segment palpe ima jedan mišić koji omogućava fleksiju sljedećeg segmenta.
U mandibulatnim usnim aparatima, labijum je četverostruka struktura, iako je formiran od dvije srasle sekundarne maksile. Može se opisati kao dno usta. S maksilama pomaže u manipulaciji hranom za vrijeme žvakanja ili drobljenja ili se, u neobičnom slučaju vretenčaste nimfe, proteže kako bi ugrabio plijen natrag u glavu, gdje ga mandibule mogu pojesti. Labijum je po strukturi sličan maksilama, ali s dodacima na obje strane stopljenim srednjom linijom, tako da čine srednju ploču. Bazni dio labija, ekvivalentan maksilarnim kardoima i koji možda uključuje i dio grudnog skeleta labijskog segmenta, naziva se postmentum. On se može podijeliti na proksimalni submentum i distalni mentum. Distalno od postmentuma i ekvivalenta sraslim maksilarnim stipulama je prementum. Prementum zatvara predornu šupljinu s leđa. Konačno, nosi četiri režnja, dvije unutrašnje glose i dvije vanjske paraglose, koji su zajedno poznate kao ligula. Jedan ili oba para režnjeva mogu biti odsutni ili mogu biti srasli u jedan srednji nastavak. Palpe nastaju sa svake strane prementuma, a često su trosegmentne.
Hipofarinks je srednji režanj neposredno iza usta, koji strši prema naprijed sa stražnje strane predusne šupljine; to je režanj nesigurnog porijekla, ali kod beskrilnih možda povezan sa donjeviličnim dijelom, kao i kod uholaža i nimfi majskih muha, kod kojih hipofarinks nosi par bočnih režnjeva, superlingvi. Šupljinu dijeli na leđnu vrećicu za hranu ili cibarij i trbušni salivarij u koji se otvara pljuvačni kanal.[1] Obično se nalazi stopljen s labijom. Većina hipofarinksa je opnasta, ali adusno lice je sklerotizirano distalno i proksimalno, sadržeći par suspenzornih sklerita koji se protežu prema gore i završavaju u bočnom zidu stomodeuma. Mišići sa polazištem na bokovima imaju hvatišta na u ovim skleritima, koji su distalno vezani za par jezičnih sklerita. Oni su zauzvrat dali hvatište zanjih antagonističke parove mišića sa polazištem na tentoriju i labijumu. Različiti mišići služe za zamah hipofarinksa prema naprijed i nazad, a u žohara još dva mišića prelaze preko hipofarinksa i proširuju pljuvačni otvor i šire salivarij.[17]
- Primjeri mandibula







Bodenje – srkanje
[uredi | uredi izvor]Usni aparat za srkanje može imati više funkcija. Neki insekti kombiniraju dijelove koji probijaju zajedno sa spužvama za upijanje koji se zatim koriste za probijanje kroz tkiva biljaka i životinja. Ženke komaraca hrane se krvlju (hemofagom, čineći ih vektorima bolesti. Usta za komarce sastoje se od proboscisa, uparenih donjih vilica i maksila. Maksile tvore iglaste strukture, nazvane stilete, koje su zatvorene labijumom. Kada komarac ugrize, maksile prodiru u kožu i učvršćuju dijelove usta, omogućavajući tako umetanje ostalih dijelova. Labij nalik ovojnici klizi natrag, a preostali dijelovi usta prolaze kroz njegov vrh i u tkivo. Zatim, kroz hipofarinks, komarac ubrizgava pljuvačku, koja sadrži antikoagulanse da zaustavi zgrušavanje krvi. I na kraju, labrum (gornja usna) koristi se za usisavanje krvi. Vrste roda Anopheles (koje najćešće napadaju balkance) karakteriziraju duge palpe (dva dijela s proširivim krajem), gotovo dosižući kraj labruma.[29]
- Primjeri usnog aparata za bodenje
-
-
-
-
Konjska muha (ženka)
-





Sifonski usni aparat
[uredi | uredi izvor]Proboscis nastaje od maksilske galeje i nalazi se u nekih insekata sa usnim aparatom za srkanje.[30] Mišići cibarija ili ždrijela snažno su razvijeni i čine pumpu. U Hemiptera i mnogih Diptera, koji se hrane tekućinama biljaka ili životinja, neke komponente usnih dijelova modificirane su za probijanje, a izdužene strukture nazivaju se stilet. Kombinirane cjevaste strukture nazivaju se proboscisi, iako se u nekim skupinama koristi specijalizirana terminologija.
Kod većine vrsta Lepidoptera sastoji se od dvije cijevi koje su međusobno povezane kukama i koje se mogu odvojiti za čišćenje. Svaka cijev je iznutra udubljena, čineći tako središnju cijev kroz koju se usisava tečnost. Usisavanje se obavlja kontrakcijom i širenjem vrećice u glavi. Proboscis je namotan ispod glave kada insekt miruje, a produžava se samo prilikom uzimanja hrane.[30] Maksilske palpe su smanjene ili čak zakržljali ostaci.[31] Uočljive su i petosegmentirane u nekim više baznim porodicama, a često su i presavijene.[8] Oblik i dimenzije proboscisa evoluirali su dajući različitim vrstama širu, a samim tim i povoljniju ishranu.[30] Postoji alometrijsko skaliranje veza između tjelesne mase leptira i dužine proboscisa [32] gdje je zanimljivo adaptivno polazište neobičnog sokolovog moljca dugog jezika Xanthopan morganii praedicta . Charles Darwin predvidio je postojanje i dužinu hobosa ovog moljca prije njegovog otkrića, na osnovu znanja o dugoprugastoj madagaskarskoj zvijezdastoj orhideji Angraecum sesquipedale.[33]
- Primjeri sifonastih usnih aparata




Upijanje
[uredi | uredi izvor]Ustni aparat insekata koji se hrane tekućinom modificirani su na razne načine kako bi se oblikovala cijev kroz koju se tekućina može uvući u usta i obično drugu kroz koju prolazi slina. Mišići cibarija ili ždrijela snažno su razvijeni da tvore svojevrsnu pumpu. U muha koje ne grizu, nema mandibula, a ostale strukture su smanjene; labijalne palpe su izmijenjene da formiraju labelum, a prisutne su i maksilarne palpe, iako ponekad kratke. U Brachycera je labelum posebno istaknut i koristi se za upijanje (spužvanje) tečne ili polutekuće hrane.[34] Labele su složene strukture od mnogih žljebova, nazvanih pseudotraheje, koje sakupljaju tečnosti. Izlučevine pljuvačke iz labela pomažu u rastvaranju i sakupljanju čestica hrane, tako da ih pseudotraheje mogu lakše uzeti; smatra se da se ovo dešava kapilarno. Tečna hrana se zatim iz pseudotraheja izvlači kroz kanal za hranu u jednjak.[35]
Usni aparati pčela su tipa za žvakanje i lizanje. Lizanje je način hranjenja u kojem se tečna ili polutekuća hrana prilijepljena za izbočeni organ ili "jezik" prenosi sa supstrata u usta. Kod medonosne pčele (Hymenoptera: Apidae: Apis mellifera ), izdužene i srasle usne glose tvore dlakavi jezik, koji je okružen maksilskom galeom i labijskim palpama, tvoreći cjevasti proboscis koji sadrži prehrambeni kanal. Pri hranjenju, jezik se uroni u nektar ili med, koji prijanja uz dlačice, a zatim se uvuči, tako da se prijanjajuća tečnost prenosi u prostor između galea i labijalnih palpi. Ovaj pokret glosa, u smjeru naprijed-natrag ponavlja se više puta. Kretanje tečnosti prema ustima očito je rezultat djelovanja cibarijske pumpe, olakšano svakim uvlačenjem jezika koji potiskuje tečnost prema kanalu za hranu.[1]
- Primjeri usnog aparata za upijanje


Toraks
[uredi | uredi izvor]Toraks insekata ima tri segmenta: protoraks, mezotoraks i metatoraks. Segment prednji, najbliži glavi, je protoraks; njegove glavne značajke su prvi par nogu i pronotum. Srednji segment je mezotoraks; njegovo glavno obilježje su drugi par nogu i prednja krila, ako ih ima. Treći, stražnji, toraksni segment, koji se naslanja na trbuh, je metatoraks, koji nosi treći par nogu i stražnja krila. Svaki segment razmeđen je međusegmentnim šavom (suturom). Svaki segment ima četiri osnovne regije. Leđna površina naziva se tergum (ili notum, da se razlikuje od trbušnog terguma).[1] Dvije bočne regije nazivaju se pleure, a trbušna naziva se sternum. Zauzvrat, notum protoraksa naziva se pronotum, notum mezotoraksa naziva se mezonotum, a notum metatoraksa je metanotum. Nastavljajući ovu logiku, tu su i mezopleura i metapleura, kao i mezosternum i metasternum.[8]
Tergalne ploče toraksa jednostavne su građe u abeskrilnih i kod mnogih nezrelih krilatih insekata, ali su različito modificirane u odraslih krilatih. Svaki pterotoraksni notum ima dva glavna odjela: prednji, krilo nosivi alinotum i stražnji, postnotum koji nosi fragmu. Fragme (jednina: fragma) su beznoge nalik pločici koji se pružaju unutra ispod antekostalnih šavova, označavajući primarne međusegmentne nabore između segmenata; fragme pružaju hvatišta za uzdužne letne mišiće. Svaki alinotum (koji se ponekad zbunjujuće naziva i "notum") može se preći šavovima koji označavaju položaj unutrašnjih ojačanih grebenovaa, a pločicu obično dijeli na tri područja: prednji preskutum, skutum i manji stražnji skutelum. Vjeruje se da lateralni pleuralni skleriti potiču od subkoksnog segmenta nogu predačkih insekata. Ovi skleriti mogu biti odvojeni, kao kod srebrnih ribica, ili stopljeni u gotovo kontinuirano sklerotizirano područje, kao kod većine krilatih insekata.[1]
Protoraks
[uredi | uredi izvor]Pronotum protoraksa može biti jednostavne građe i malen u usporedbi s ostalim notumima, ali u tvrdokrilaca, mantida i nekih vrsta Orthoptera, pronotum je proširen, a u žohara čini štit koji pokriva dio glava i mezotoraks.[1][8]
Pterotoraks
[uredi | uredi izvor]Budući da mezotoraks i metatoraks drže krila, oni imaju kombinirano ime zvano pterotoraks (pteron = krilo). Prednje krilo, koje nosi različita imena u različitim redovima (npr. tegmina u Orthoptera i elitra u Coleoptera), nastaje između mezonotuma i mezopleurona, a zadnje krilo zgloblljava se između metanotuma i metapleurona. Noge nastaju iz mezopleurona i metapleure. Mezotoraks i metatoraks imaju pleursku suturu (mezonotumskog i metapleurskog šava) koji ide od osnove krila do nožne kokse. Sklerit ispred pleurskog šava naziva se episternum (serijski, mezepisternum i metepisternum). Sklerit koji je straga od šava je epimiron (serijski, mesepimiron i metepimiron). Spirakule, vanjski organi respiratornog sistema, nalaze se na pterotoraksu, obično između pro– i mezopleorona, kao i između mezo– i metapleurona.[8]
Ventralna površina ili sternum slijede istu konvenciju, s prosternumom ispod protoraksa, mezosternumom ispod mezotoraksa i metasternumom ispod metatoraksa. Notum, pleura i grudna ploča svakog segmenta imaju niz različitih skleritisa i šavova, koji se uvelik razlikuju od reda do reda, i o njima se neće detaljno raspravljati u ovom odjeljku.[8]
Krila
[uredi | uredi izvor]Većina filogenetski naprednih insekata ima dva para krila koja se nalaze na drugom i trećem toraksnom segmentu.[1] Insekti su jedini beskičmenjaci koji su razvili sposobnost leta, a to je imalo važnu ulogu u njihovom uspjehu. Let insekata nije dobro razumljiv, oslanjajući se uveliko na turbulentne aerodinamičke efekte. Primitivne skupine insekata koriste mišiće koji djeluju direktno na strukturu krila. Naprednije grupe, koje čine Neoptera, imaju sklopiva krila, a mišići djeluju na zid grudnog koša i indirektno pokreću krila.[1] Ovi mišići mogu se kontraktirati više puta za svaki pojedinačni nervni impuls, omogućavajući krilima da pokreću brže nego što bi to obično bilo moguće.
Let insekata može biti izuzetno brz, upravljiv i svestran, moguće zbog promjenjivog oblika, izvanredne kontrole i promjenjivog kretanja krila. Redovi insekata imaju različite mehanizme leta; naprimjer, let leptira može se objasniti korištenjem teorije stabilnog stanja, netranzitorijske aerodinamike i teorije tankog aeroprofilnosti.

Unutrašnjost
[uredi | uredi izvor]Svako krilo sastoji se od tanke opne poduprte sistemom „vena“ ili „nervature“. Membrana je sastavljena od dva sloja prekrivača koji su tijesno postavljeni, dok vene nastaju tamo gdje dva sloja ostaju odvojena, a kutikula može biti gušća i jače sklerotizirana. Unutar svake od glavnih vena nalazi se živac i traheja, a budući da su šupljine vena povezane sa hemocelom, hemolimfa može teći u krila.[26] Kako krilo se razvija, leđni i trbušni integumentni slojevi postaju usko postavljeni na većini svog područja, čineći membranu krila. Preostala područja čine kanale, buduće vene, u kojima se mogu pojaviti živci i traheje. Kutikula koja okružuje vene postaje zadebljala i jače se sklerotizira, kako bi pružila čvrstoću i krutost krilu. Na krilima se mogu pojaviti dlake dvije vrste: mikrotrihe, koje su male i nepravilno raspršena, i makrotrihe, koje su veće, udubljene i mogu biti ograničene na vene. Ljuske Lepidoptera i Trichoptera su visoko modificirana makrotrihe.[27]
Vene
[uredi | uredi izvor]
Kod nekih vrlo malih insekata, venacija može biti znatno smanjena. Naprimjer, halkidne ose imaju samo subkoste i dio radijusa. Suprotno tome, do povećanja venacije može doći razgranavanjem postojećih vena radi stvaranja pomoćnih vena ili razvojem dodatnih, interkalarnih vena između izvornih, kao u krilima Orthoptera (skakavaca i cvrčaka). Veliki broj ukrštenih vena prisutan je kod nekih insekata i oni mogu formirati mrežu kao u krilima Odonata i u osnovi prednjih krila Tettigonioidea i Acridoidea .[26]
Arhediktion je ime dato hipotetskoj shemi venacije krila koja je predložena za prvog krilatog insekta. Zasnovan je na kombinaciji spekulacija i fosilnih podataka. Budući da se vjeruje da su svi krilati insekti evoluirali od zajedničkog pretka, arhedikton predstavlja "predložak" koji je prirodna selekcija modificirala (i pojednostavila) tokom 200 miliona godina. Prema postojećoj dogmi, zasvođeni je šest do osam uzdužnih vena. Te vene (i njihove grane) nazvane su prema sistemu koji su osmislili John Comstock i George Needham – Comstock-Needhamov sistem:[36]
- Kosta (C) – vodeća ivica krila;
- Subkosta (Sc) – druga uzdužna vena (iza koste), tipično nerazgranata;
- Radijus (R) – treća uzdužna vena, jedna do pet grana doseže obod krila;
- Medija (M) – četvrta uzdužna vena, jedan do četiri kraka dosežu ivicu krila;
- Kubitus (Cu) – peta uzdužna vena, sa jedan do tri kraka koji dosežu ivicu krila;
- Analne veine (A1, A2, A3) – nerazgranate vene iza lakata.
Kosta (C) je vodeća rubna vena kod većine insekata, iako se mala vena, prekosta, ponekad nalazi iznad koste. Kod gotovo svih postojećih insektata,[1], prekosta je srasla s kostom; kosta se rijetko ikad grana, jer je na prednjem rubu, koji je u osnovi povezan s humeralnom pločom. Traheja rebrene vene možda je grana subkostalne traheje. Nakon koste smještena je treća vena, potkosta, koja se grana u dvije odvojene vene: prednju i stražnju. Baza podkoste povezana je s distalnim krajem vrata prve aksilarne koste. Četvrta vena je radijus koji je razgranat u pet zasebnih vena. Radijus je općenito najjača vena krila. Prema sredini krila račva se u prvi nepodijeljeni krak (R1) i drugi krak, nazvan radijalni sektor (Ra), koji se dihotomno dijeli na četiri distalna kraka (R2, R3, R4, R5). U osnovi, radijus je fleksibilno sjedinjen s prednjim krajem druge aksilarne (2Ax).[37]
Peta vena krila je medija. U arhetipskom uzorku (A), medija se račva u dvije glavne grane, prednju (MA), koja se dijeli na dvije distalne grane (MA1, MA2) i srednji sektor, ili stražnju (MP), koji ima četiri terminalne grane (M1, M2, M3, M4). U većine modernih insekata, prednji dio medije je izgubljen, a uobičajena "medija" je stražnja četverorazgranata medija sa zajedničkom bazalnom drškom. U Ephemeroptera, prema sadašnjim tumačenjima venacije krila, zadržane su obje grane medija, dok je u Odonata postojana medija primitivna prednja grana. Stabljika medija često je sjedinjena s radijusom, ali kada se javlja kao posebna vena, njena baza je povezana s distalnom pločicom medije (m') ili se kontinuirano sklerotizira s ovom drugom. Kubitus, šesta vena krila, prvenstveno je dvokraka. Primarno račvanje odvija se blizu dna krila, formirajući dva glavna kraka (Cu1, Cu2). Prednja grana može se dijeliti na niz sekundarnih grana, ali obično se račva na dvije distalne grane. Comstock i Needham zamijenili su drugu granu kubitusa (Cu2) u Hymenoptera, Trichoptera i Lepidoptera za prvu analnu. Proksimalno, glavno stablo kubita povezano je s distalnom srednjom pločom (m') osnove krila.[37]
Postkubitus (Pcu) je prva analna vena u sistemu Comstocka i Needhama. Međutim, on ima status neovisne krilne vene i trebao bi biti prepoznat kao takav. U nimfalnim krilima, traheja nastaje između kubitusne traheje i skupine vanalnih traheja. U zrelim krilima generaliziranih insekata, postkubitus je uvijek povezan proksimalno s kubitusom i nikada nije usko povezan sa fleksornim skleritom (3Ax) osnove krila. U Neuroptera, Mecoptera i Trichoptera, postkubitus je možda usko povezan s vanalnim venama, ali njegova baza je uvijek slobodna od potonjih. Postkubitus je obično nerazgranat; primitivno je dvokraka vena. Vanalne vene (lV do nV) su analne vene koje su odmah povezane sa trećom aksilarnom paletom i na koje izravno utiče kretanje ovog sklerita koji dovodi do savijanja krila. Brojne, vanalne vene variraju od jedne do 12, u zavisnosti od širenja vanalnog područja krila. Vanalne traheje obično nastaju iz zajedničke trahejne stapke kod nimfnih insekata, a vene se smatraju granama jedne analne vene. Distalno, vanalne vene su ili jednostavne ili razgranate. Jugalna vena (J) jugalni režanj krila često su sa mrežom nepravilnih vena ili je možda u potpunosti opnasta; ponekad sadrži jednu ili dvije različite, male vene, prvu jugalnu venu ili lučnu venu i drugu jugalnu venu ili venu kardinalis (2J).
- C-Sc Poprečne vene – između kosta i subkosta;
- R poprečne vene – između susjednih grana;
- R-M poprečne vene – između radijusa i medije;
- M-Cu poprečne vene – između medije i kubitusa.
Sve vene krila podložne su sekundarnom račvanju i spajanju poprečnim venama. U nekim redovima insekata, poprečne vene su toliko brojne, da čitav venacijski obrazac postaje uska mreža razgranatih i poprečnih vena. Međutim, na određenim lokacijama, obično se javlja određeni broj unakrsnih vena. Stalnije poprečne vene su humeralna poprečna vena (h). između koste i podkoste, radijalna poprečna vena (r) između R i prve račve Rs, sektorska poprečna vena između dvije račve R8, srednja poprečna vena (mm) između M2 i M3, i mediokubitusna poprečna vena (m-cu) medije i kubita.[37]
Vene krila insekata karakteriziraju konveksno-udubljeni položaj, kakav se vidi kod muha (tj. udubljenje je "dolje", a konveksno "gore"), koje se redovito izmjenjuju i grananjem; kad god se vena račva, između dvije grane uvijek postoji interpolirana vena suprotnog položaja. Konkavna vena će se račvati u dvije konkavne vene (s interpoliranom venom koja je konveksna) i očuvana je redovita varijacija vena. .[38] Čini se da krilne vene dobijaju talasast obrazac, prema tome imaju li tendenciju da se preklope gore ili dolje kada je krilo opušteno. Bazne osovine vena su konveksne, ali svaka se račva distalno u prednju konveksnu i stražnju konkavnu granu. Dakle, kosta i podkosta smatraju se konveksnim i konkavnim granama prve primarne vene, Rs je konkavna grana radijusa, a stražnja medija je konkavni krak medija, Cu1 i Cu2 su konveksne i konkavne, dok su primitivni postkubitus, a prve vanalne imaju po jedan prednji konveksni i stražnji konkavni krak. Konveksna ili udubljena priroda vena korištena je kao dokaz pri određivanju identiteta postojanih distalnih grana vena modernih insekata, ali nije dokazano da je konzistentna za sva krila.[26][37]
Polja
[uredi | uredi izvor]
Područja krila su razdvojena i podijeljena preklopnim linijama , duž kojih se krila mogu preklopiti i linijama savijanja, koje se savijaju tokom leta. Između linija savijanja i preklapanja, osnovna razlika često je zamagljena, jer linije preklapanja mogu omogućiti određenu fleksibilnost ili obrnuto. Dvije konstante, koje se nalaze u gotovo svim krilima insekata, su klaval (linija savijanja) i jugalalni nabor (ili linija nabora), čineći promjenljive i nejasne granice. Nabori krila mogu se vrlo zakomplicirati, pri čemu se poprečno nabiranje može dogoditi u stražnjim krilima Dermaptera i Coleoptera, a kod nekih insekata područje anusa može se sklopiti poput lepeze.[1]:41–42 Četiri različita polja na krilima insekata su:
- Remigij
- Analno područje (vanus)
- Jugalno područje
- Pazušno područje
- Alula
Većina običnih i unakrsnih vena javlja se u prednjem području remigija, koji je odgovoran za veći dio leta, a pokreću ga toraksni mišići. Stražnji dio remigija ponekad se naziva klavus; dva druga zadnja polja su analno i jugalno područje.[1] Kada vanalni nabor ima uobičajeni položaj ispred grupe analnih vena, remigij sadrži rebrene, subkostalne, radijalne, medijalne, kubitalne i postkubitalne vene. U flektiranom krilu remigij se okreće straga na fleksibilnoj baznoj vezi radijusa s drugom aksilarnom venom, a osnova mediokubitalnog polja je medijalno presavijena na aksilarnom području duž plica basalis (bf), između srednjih ploča (m, m') baze krila.[37]
Vanus je omeđen vanalnim naborom, koji se obično javlja između postkubitusa i prve vanalne vene. U Orthoptera obično ima ovakav položaj. Međutim, u prednjem krilu porodice Blattidae, jedini nabor u ovom dijelu krila je neposredno prije postkubitusa. U Plecoptera, vanalni nabor je straga od postkubitusa, ali proksimalno prelazi dno prve vanalne vene. U cikada, vanalni nabor je neposredno iza prve vanalne vene (lV). Ove male varijacije u stvarnom položaju vanalnog nabora, međutim, ne utiču na jedinstvo djelovanja vanalnih vena, kontroliranih fleksorskim skleritom (3Ax), u savijanju krila. U stražnjim krilima većine Orthoptera, sekundarna vena djeli bokove u vanjskom naboru. Vanus je obično trouglast, a vene se tipski šire od treće aksilarne kostalne poput rebara ventilatora. Neke od vanalnih vena mogu biti razgranate, a sekundarne se mogu izmjenjivati s primarnim venama. Vanalna regija obično je najbolje razvijena u stražnjem krilu, u kojoj se može proširiti tako da tvori potpornu površinu, kao u Plecoptera i Orthoptera. Velika proširenja stražnjih krila u porodici Acrididae nalik lepezi očito su vanjske regije, jer su sve njihove vene oslonjene na treće pazušne sklerite na krilnim osnovama, iako Martynov (1925) većinu lepeznih područja u Acrididae pripisuje jugalima krila. Pravi jugulum njihovog krila predstavljen je samo malim membranskim (Ju) mesadom posljednje vanalne vene. Jugum je jače razvijen u nekih drugih ortotera, kao u mantida. U većine viših insekata uskih krila vannus se smanjuje, a vanna nabora se gubi, ali čak i u takvim slučajevima, savijeno krilo može se saviti duž linije između postkubita i prve vanjske vene,[37]
Jugalna regija, ili neala, je područje krila koje je (obično malo) opnasto područje proksimalno od dna vanusa, ojačano s nekoliko malih, nepravilnih zadebljanja nalik venama; ali kada je dobro razvijen, to je poseban dio krila i može sadržavati jednu ili dvije vratne vene. Kada se područje jugala prednjeg krila razvije kao slobodni režanj, izbacuje se ispod humeralnog ugla zadnjeg krila i na taj način služi za spajanje dvaju krila. U grupi Jugatae Lepidoptera postoji dugačak režanj poput prsta. Jugalska regija nazvana je neala ("novo krilo") jer je očito sekundarni i nedavno razvijeni dio krila.[37]
Pomoćno područje koje sadrži aksilarne sklerite ima, općenito, oblik skleritnog trougla. Osnova trougla (a–b) je šarka krila s tijelom; vrh (c) je distalni kraj trećeg aksilarnog sklerita; duža strana je sprijeda na vrhu. Tačka d na prednjoj strani trougla označava zglob radijalne vene s drugim aksilarnim skleritom. Linija između d i c je plica basalis (bf), odnosno nabor krila u dnu mediokubitusnog polja.[37]
U stražnjem uglu osnove krila kod nekih dvokrilnih Diptera nalazi se par opnastih režnjeva (skvame ili kaliptere) poznate kao alule. Alula je dobro razvijena kod kućne muhe. Vanjska skvama (c) nastaje iz osnove krila, iza trećeg aksilarnog sklerita (3Ax) i očito predstavlja jugalusni režanj ostalih insekata (A, D); veća unutrašnja skvama (d) nastaje iz stražnjeg zglobnog ruba terguma, segmenta koji nosi krilo i čini zaštitnu nadstrešnicu poput kapuljače preko utora. U savijenom krilu vanjska skvama alule okrenuta je naopako iznad unutrašnje skvame, na koju potonji ne utiče kretanjem krila. Kod mnogih Diptera duboki rez analnog područja krilne membrane iza pojedinačne vanalne vene postavlja proksimalni krilni režanj distalno od vanjske skvame alule.[37]
Zglobovi
[uredi | uredi izvor]
Različiti pokreti krila, posebno kod insekata koji savijaju krila vodoravno preko leđa kada miruju, zahtijevaju složeniju zglobnu strukturu u osnovi krila od pukog zgloba krila s tijelom. Svako je krilo za tijelo pričvršćeno membranskim baznim područjem, ali zglobna opna sadrži niz malih zglobnih sklerita, zajednički poznatih kao pteralije. Pteralije uključuju prednju humeralnu ploču u dnu kostalne vene, skupinu aksilarnih (Ax) povezanih sa subkostalnim, radijalnim i vanalnim venama i dvije manje određene srednje srednje ploče (m, m ') u dnu mediokubitusnog područja. Aksilari su posebno razvijeni samo kod insekata koji savijaju krila, gdje imaju mehanizam fleksije krila kojim upravlja mišić fleksor koji polazi na pleuronu. Karakteristika baze krila je i mali režanj na prednjoj margini zglobnog područja, proksimalno od humeralne ploče, koji je na prednjem krilu nekih insekata razvijen u veliku, ravnu klapnu nalik skali, tegulu koja se preklapa u osnovi krila. Na stražnjoj strani, zglobna opna često formira obilni režanj između krila i tijela, a rub joj je uglavnom zadebljan i rebrast, dajući izgled ligamenta, takozvane aksilarne vrpce, kontinuirane mezalno sa stražnjim rubnim skutelusnim naborom, tergitna ploča koja nosi krilo.[37]
Zglobni skleriti ili pteralije, baze krila insekata koji se savijaju i njihovi odnosi prema tijelu i krilnim žilama, shematski su prikazani, kako slijedi:
- Humeralne ploče ;
- Prva pazušna (aksilarna);
- Druga pazušna;
- Treća pazušna;
- Četvrta pazišna;
- Medijalne ploče (m, m')
Humeralna ploča obično je mali sklerit na prednjem rubu osnove krila, pomičan i zglobno povezan s dnom rebrene vene. Kod Odonata su humeralne ploče uveliko povećane,[37] s dva mišića sa polazištem iz episternuma i hvatištem na humeralnm pločama i dva na rubu epimerona na pazušnoj (aksilarnoj) ploči.[26]
Prvi aksilarni sklerit (lAx) je prednja zglobna ploča baze krila. Njegov prednji dio je oslonjen na prednji otvor notusnog krila terguma (ANP); stražnji dio mu se zglobljava s tergitnom ivicom. Prednji kraj skleritita uglavnom je u obliku vitkog dodatka čiji je vrh (e) uvijek povezan s bazom subkostalne vene (Sc), iako nije sjedinjen s ovom drugom. Tijelo skleritita zglobljava se bočno s drugom aksilarnom. Drugi aksilarni sklerit (2Ax) je varijabilnijeg oblika od prvog, ali njegovi mehanički odnosi nisu ništa manje određeni. Koso je spojena na vanjski rub tijela prve pazušne jame, a radijalna vena (R) uvijek je fleksibilno pričvršćena za svoj prednji kraj (d). Druga aksilarna predstavlja i leđnu i trbušnu sklerotizaciju u osnovi krila; njegova trbušna površina počiva na potpornom krilu pleurona. Stoga je drugi aksilarni stožerni sklerit dna krila i on posebno manipulira radijalnom venom.[37]
Treći aksilarni (pazušni) sklerit (3Ax) leži u stražnjem dijelu zglobne regije krila. Njegov oblik je vrlo varijabilan i često je nepravilan, ali treća aksilarna je sklerit na kojem je hvatište za mišiće fleksore krila (D). Mezalno, zglobljava se sprijeda (f) sa stražnjim krajem druge aksilarne ploče, a straga (b) sa stražnjim krilnim otvorom terguma (PNP), ili sa malom četvrtom aksilarnom kada je ova prisutna. Distalno, treća aksilarna ploča produžava se u nastavak koji je uvijek povezan sa bazama grupe vena u analnom području krila, ovdje zvanom vanalne vene (V). Stoga je treća pazušna ploča obično stražnja zglobna ploča baze krila i aktivni je sklerit mehanizma savijača, koji direktno manipulira vanalnim venama. Kontrakcija mišića fleksora (D) okreće treću aksilarnu venuu na svojim mezalnim zglobovima (b, f) i time podiže njegov distalni krak; ovaj pokret izaziva savijanje krila. Četvrti aksilarni sklerit nije stalni element osnove krila. Kad je prisutan, to je obično mala ploča koja intervenira između treće aksilarne ploče i stražnjeg notumskog otvora i vjerojatno je odvojeni dio potonjeg.[37]
Medijanske ploče (m, m ') također su skleriti, koji nisu toliko definitivno diferencirani kao specifične ploče, kao što su tri glavne aksilarne ploče, ali su važni elementi aparata za savijanje. Leže u srednjem području baze krila, distalno od druge i treće aksilarne šare, a međusobno su odvojene kosom linijom (bf), koja stvara istaknuti konveksni nabor tokom savijanja krila. Proksimalna ploča (m) obično je pričvršćena za distalni krak treće aksilarne i možda bi je trebalo smatrati dijelom potonje. Distalna ploča (m ') manje je stalno prisutna kao izraziti sklerit, a može biti predstavljena općom sklerotizacijom osnove mediokubitusnog polja krila. Kada se vene ovog područja u osnovi razlikuju, povezane su s vanjskom medijanskom pločom.[37]
Spajanje, preklapanje i druga obilježja
[uredi | uredi izvor]Kod mnogih vrsta insekata prednje i zadnje krilo su povezana, što poboljšava aerodinamičku efikasnost leta. Najčešći mehanizam spajanja (npr. Hymenoptera i Trichoptera) je niz malih kukica na prednjem rubu zadnjeg krila, ili "hamuli", koji se zaključavaju na prednjem krilu, držeći ih zajedno (hamulatna spojnica). U nekih drugih vrsta insekata (npr. Mecoptera, Lepidoptera i nekih Trichoptera jugalusni prednji dio prednje krilice pokriva dio zadnjeg krila (juglska spojnica) ili se margine prednjeg i stražnjeg krila široko preklapaju (ampleksiformna spojnica), ili čekinje zadnjih krila, ili frenulum, zakače se ispod potporne konstrukcije ili mreže na prednjem krilu ===== Coupling, folding, and other features.[1]:43
Kada miruju, krila se kod većine insekata drže preko leđa, što može uključivati uzdužno presavijanje membrane krila, a ponekad i poprečno savijanje. Preklapanje se ponekad može dogoditi duž linija savijanja. Iako linije nabora mogu biti poprečne, kao na stražnjim krilima tvrdokrilaca, obično su radijalne u odnosu na podnožje krila, omogućavajući da se susjedni dijelovi krila preklope jedan preko drugog ili ispod njih. Najčešća linija nabora je jugalni nabor, smješten odmah iza treće analne vene,[27], iako većina Neoptera ima jugalni nabor odmah iza vene 3A, na prednjim krilima. Ponekad je prisutan i na stražnjim krilima. Tamo gdje je analno područje stražnjeg krila veliko, kao kod Orthoptera i Blattodea, cijeli ovaj dio može se sklopiti ispod prednjeg dijela krila, duž vanjskog nabora malo pozadi klavalne brazde. Pored toga, kod ortotera i blatodea, analno područje je uređeno poput lepeze duž vena, pri čemu su analne vene konveksne, na grebenima nabora, a pomoćne vene udubljene. Dok su klavalna brazda i jugalni nabor vjerovatno homologni kod različitih vrsta, vanjski nabor, kod različitih taksona, varira u položaju. Preklapanje pokreće mišić sa polazištem na pleuronu i hvatištem na trećem aksilarnom skleritu, na takav način da se, kada se kontrahira, sklerit okreće oko svojih tačaka artikulacije sa stražnjim notusnim otvorom i drugim aksilarnim skleritom.[26]
Noge
[uredi | uredi izvor]Tipski i uobičajeni segmenti noge insekta podijeljeni su na koksu, jedan trohanter, butni dio, potkoljenični, tarzus i pretarzus. Koksa u svom više simetričnom obliku ima oblik kratkog cilindra ili krnjeg konusa, iako je obično jajasta i može biti gotovo sferna. Proksimalni kraj cokse opasan je submarginalnim bazikostalnim šavom koji iznutra formira greben ili bazikostu i postavlja rubnu prirubnicu, koksomarginal ili bazikoksite . Bazikosta jača bazu korijena i obično je uvećan na vanjskom zidu kako bi se prihvatili mišići; na mezalnoj polovini, koksa je, međutim, obično slaba i često se ulijeva u koksni rub.
Trohanterskii mišići čije je polazište coksa uvijek su pričvršćeni distalno od bazikoste. Koksa je za tijelo pričvršćeno zglobnom opnom, coxal corium, koje okružuje njegovu bazu. Ova dva zglobljavanja možda su primarne leđne i trbušne zglobne tačke subkokso-koksne šarke.Pored toga, insektska koksa često ima prednje zglobljavanje sa prednjim, trbušnim krajem trohantera, ali artikulacija trohanteraa ne koegzistira sa artikulacijom grudne koste. Pleuralna zglobna površina kokse leži na mezalnom pregibu koksnog zida. Ako je koksa pokretna samo na pleuralnoj artikulaciji, zglobna površina koksala obično se savija do dovoljne dubine da pruži podršku mišićima odmicačima sa hvatištem na vanjski obod baze koksala. Distalno koksa nosi prednju i stražnju artikulaciju sa trohanterom. Vanjski zid kokse često je obilježen šavom koji se proteže od baze do prednje trohanterske artikulacije. Kod nekih insekata, koksni šav je u liniji s pleurnim šavom, a u takvim slučajevima čini se da je koksa podijeljena na dva dijela, koji odgovaraju episternumu i epimeronu pleurona. Mnogi insekti nemaju koksni šav.[37]
Pregib koksnog zida, koji nosi pleurnu zglobnu površinu dijeli bočni zid bazikoksita na predzglobni i postzglobni dio, a ta dva područja često se pojavljuju kao dva rubna režnja na osnovi koksa. Stražnji režanj je obično veći i naziva se meron. Meron se može uvećati produžetkom distalno u stražnjem zidu koksa; u Neuroptera, Mecoptera, Trichoptera i Lepidoptera. meron je toliko velik da se čini da je koksa podijeljena na prednji dio, takozvani "coxa genuina" i meron, ali meron nikada ne uključuje područje stražnja trohanterska artikulacija, a žlijeb koji je ograničava uvijek je dio bazikostnog šava. Koksa s uvećanim meronom ima izgled sličan onome podijeljenom koksnim šavom koji je u liniju s pleurnim šavom, ali ta su dva stanja u osnovi prilično različita i ne bi ih trebalo miješati. Meron dostiže krajnje odstupanje od uobičajenog stanja Diptera. U nekih općenitijih muha, kao u Tipulidae, meron srednje noge izgleda kao veliki režanj kokse koji strši prema gore i straga iz baze koksala; kod viših članova reda postaje potpuno odvojen od kokse i formira ploču bočnog zida mezotoraksa.[37]
Trohanter je bazalni segment telopodita; to je uvijek mali segment u nozi insekta, koji se slobodno pomiče vodoravnim šarkama na korijenu, ali je više ili manje fiksiran na dnu bedrene koste. Kada se pomiče na bedru, trohantero-femoralna šarka je obično vertikalna ili kosa u vertikalnoj ravni, dajući lagano kretanje i smanjenje na zglobu, iako je prisutan samo reduktor. U Odonata, i nimfe i odraslih, postoje dva trohanterna segmenta, ali nisu pokretni jedan na drugom; drugi sadrži redukcijski mišić bedrene kosti. Uobičajeni pojedinačni trohanterni segment insekata, prema tome, vjerovatno predstavlja dva trohantera drugih člankonožaca koji su stopljeni u jedan prividni segment, jer nije vjerovatno da je primarni zglob koksotrohantera izgubljen s noge. U nekim od Hymenoptera, bazna podjenica bedrene koste simulira drugi trohanter, ali hvatište reduktorskog mišića na njegovu bazu potvrđuje da pripada segmentu femoralne koste, jer, kako je prikazano na odonatnoj nozi, reduktor ima polazište iz pravog drugog trohantera.[37]:165
Femur je treći segment noge insekta i obično je najduži i najjači dio udova, ali varira u veličini, od ogromne zadnje bedrene kosti koja skače kod Orthoptera, do vrlo malog segmenta kakav je prisutan u mnogih larvenih oblika. Volumen femura u pravilu je u korelaciji s veličinom tibijskih mišića koji se u njemu nalaze, ali ponekad se povećava i modificira u oblik za druge svrhe, osim za hvatište tibijskih mišića. Tibija je karakterističan tanak segment kod odraslih insekata, samo malo kraći od femura ili kombiniranog femura i trohantera. Njen proksimalni kraj čini manje ili više različitu glavu, savijenu prema femuru, uređaj koji omogućava da se potkoljenica savija blizu donje površine bedra.
Termini profemur, mezofemur i metafemur odnose se na femure prednjih, srednjih i stražnjih nogu insekata.[39] Slično se protibija, mezotibija i metatibija odnose na tibije prednjih, srednjih i stražnjih nogu.[40]
Tarzus (stopalo) insekata odgovara pretposljednjem segmentu uopćenih udova člankonožaca, a to je segment nazvan propodit u Crustacea. U odraslih insekata obično je podijeljen na dva do pet podsegmenata ili tarzomera, ali u (beskrilnih) Protutura, nekih Collembola i većini holometabolnih larvi insekata, on zadržava primitivni oblik jednostavnog segmenta. Podsegmenti odraslog insekta tarzusa su obično su slobodno pomični jedan preko drugog, pomoću pregibnih spojnih membrana, ali tarzus nikada nema unutarnje mišiće. Tarzus odraslih krilatih insekata, koji imaju manje od pet podsegmenata, vjerovatno je specijaliziran gubitkom jednog ili više ili fuzijom susjednih podsegmenata. U tarzusu porodice Acrididae, dugi bazni dio očito je sastavljen od tri sjedinjene tarzomere, ostavljajući četvrtu i petu. Bazna tarzomera je ponekad vidljivo povećana i razlikuje se kao bazitarzus. Na donjim površinama tarzusnih podsegmenata, kod određenih pravokrilaca, nalaze se mali jastučići, tarzusin pulvili ili euplantule. Tarzus se povremeno stopi s tibijom kod larvi, čineći tibiotarzusni segment; u nekim se slučajevima čini da se eliminira ili svodi na rudiment između tibije i pretarzusa.[37]
Femur i tibija su uglavnom najduži segmenti nogu, ali varijacije u dužini i robusnosti svakog segmenta odnose se na njihove funkcije. Naprimjer, gresorijski i kursorijski, odnosno insekti koji hodaju i trče, obično imaju dobro razvijene femure i tibije na svim nogama, dok su skakajući (saltatorijski) insekti poput skakavaca nesrazmjerno razvili metafemure i metatibije. U vodenih tvrdokrilaca (Coleoptera) i buha (Hemiptera), tibija i / ili tarzusi jednog ili više parova nogu obično su modificirani za plivanje (u prirodi) s resama dugih, vitkih dlaka. Mnogi insekti koji obitavaju u tlu, poput cvrčaka (Orthoptera: Gryllotalpidae), cifadnih nimfi (Hemiptera: Cicadidae) i skarabeja (Scarabaeidae), imaju tibije prednjih nogu ( protibije) uvećane i modificirane za kopanje (fosorijalne), dok su prednje noge nekih grabežljivih insekata, poput mantispidnih (Neuroptera) i mantida (Mantodea), specijalizirane za oduzimanje plijena ili grabežljivca. Tibija i bazne tarzomere svake zadnje noge medonosnih pčela modificirane su za sakupljanje i transport polena.[26]
Trbuh ili zadak
[uredi | uredi izvor]Plan građe abdomena odraslog insekta obično se sastoji od 11-12 segmenata, slabije sklerotiziranih od glave ili toraksa. Svaki segment predstavljen je sklerotiziranim tergom, sternumom i možda pleuritom. Tergiti su međusobno odvojene membranom, kao i od susjedne sterne ili pleure. Spirakule se nalaze u pleurnom području. Varijacija ovog planova uključuje spajanje tergita ili tergita i sternuma, da bi se stvorili kontinuirani leđni ili trbušni štit ili konusna cijev. Neki insekti imaju sklerit u pleurnom području koji se naziva laterotergit. Ventralni skleriti se ponekad nazivaju laterosterniti. Tokom embrionske faze mnogih insekata i postembrionske faze primitivnih insekata, prisutno je 11 trbušnih segmenata. U modernih insekata postoji tendencija ka smanjenju broja trbušnih segmenata, ali tokom embriogeneze zadržava se primitivni broj 11. Varijacija broja trbušnih segmenata je značajna. Ako se Apterygota smatra indikativnim za plan građe pterigota, vlada zabuna: odrasli beskrilni Protura imaju 12 segmenata, Collembola ima 6. Pravokrilska porodica Acrididae ima 11 segmenata, a fosilni primjerak Zoraptere ima 10-segmentni trbuh.[8]
Općenito, prvih sedam trbušnih segmenata odraslih (pregenitalni segmenti) su slične građe i nemaju dodataka. Međutim, apterigoti (beskrilni) i mnogi nezreli vodeni krilati insekti imaju trbušne dodatke. Aterigote posjeduju par načina; rudimentarni dodaci koji su serijski homologni s distalnim dijelom toraksnih nogu. I, mezalno, jedan ili dva para izbočenih (ili eksertilnih) vezikula na barem nekim trbušnim segmentima. Ove vezikule su izvedene iz endama kokse i trohanterala (unutrašnji prstenasti režnjevi) trbušnih dodataka predaka. Vodene larve i nimfe mogu imati škrge bočno na nekim do većine trbušnih segmenata.[1] Ostali trbušni segmenti sastoje se od reproduktivnog i analnog dijela.
Analno-genitalni dio trbuha, poznat kao terminalije, sastoji se uglavnom od 8 ili 9 segmenata do trbušnog vrha. Segmenti 8 i 9 nose genitalije; segment 10 vidljiv je kao cjelovit segment kod mnogih "nižih" insekata, ali uvijek nema dodataka; mali segment 11 predstavljen je leđnim epiproktom i parom ventralnih paraprokata, izvedenih iz sternuma. Par dodataka, cerci, artikulira se bočno na segmentu 11; obično su prstenasti i nitasti, ali su i modificirane (npr. kliješta uholadža) ili smanjeni u različitim redovima insekata. Prstenasti repni filament, srednji dodatak dorsalis, nastaje od vrha epiprokta kod apterigota, većine muharica (Ephemeroptera) i nekoliko fosilnih insekata. Slična struktura kod nimfskih muha (Plecoptera) je neizvjesne homologije. Ovi terminalni trbušni segmenti imaju izlučujuću i senzornu funkciju kod svih insekata, ali kod odraslih imaju i dodatnu, reproduktivnu funkciju.[1]
Vanjske genitalije
[uredi | uredi izvor]
Organi koji posebno služe za parenje i polaganje jaja, skupno su poznati kao vanjske genitalije, iako mogu biti uglavnom unutrašnje. Komponente vanjskih genitalija insekata vrlo su raznolike u obliku i često imaju znatnu taksonomsku vrijednost, posebno među vrstama koje se u ostalom dijelu čine strukturno slične. Muške vanjske genitalije široko se koriste za pomoć u razlikovanju vrsta, dok su ženske vanjske genitalije jednostavnije i manje raznolike.
Terminali odraslih ženskih insekata uključuju unutrašnje strukture za prihvat muškog kopulacijskog organa i njegovih spermatozoida, te vanjske strukture koje se koriste za jaja (polaganje jaja; odjeljak 5.8). Većina ženki insekata ima cijev za polaganje ili ovipoziciju jaja; nema je kod termita, parazitskih uši, mnogih Plecoptera i većine Ephemeroptera. Ovipozitori imaju dva oblika:
- pravi ili apendikulski, nastal od dodataka trbušnih segmenata 8 i 9;
- supstitucijski, sastavljen od rastezljivih stražnjih trbušnih segmenata.
- Ostali dodaci
Unutrašnjost
[uredi | uredi izvor]Nervni sistem
[uredi | uredi izvor]Nervni sistem insekta može se podijeliti na mozak i ventralnu nervnu vrpcu. Glavena kapsula sastoji se od šest sraslih segmenata, svaki sa parom ganglia, ili skupom nervnih ćelija izvan mozga. Prva tri para ganglija srasla su u mozak, dok su sljedeća tri para srasla u strukturu od tri para ganglija ispod jednjaka, zvanih podjednjačka ganglija..[1]
Toraksni segmenti imaju po jednu gangliju sa svake strane, koje su povezane u par, po jedan par po segmentu. Ovakav raspored vidi se i na trbuhu, ali samo u prvih osam segmenata. Mnoge vrste insekata su smanjile broj ganglija zbog fuzije ili redukcije.[41] Neki žohari imaju samo šest ganglija u abdomenu, dok osa Vespa crabro ima samo dva u toraksu i tri u trbuhu. Neki insekti, poput kućne muhe Musca domestica, imaju sve tjelesne ganglije stopljene u jednu veliku toraksnu gangliju.
Barem nekoliko insekata ima nociceptore, ćelije koje otkrivaju i prenose osjećaje bola.[42] Ovo je otkriveno 2003., proučavanjem varijacija u reakcijama larvi obične voćne mušice roda Drosophila na dodir zagrijane i negrijane sonde. Larve su na dodir zagrijane sonde reagirale stereotipnim ponašanjem kotrljanja kojeg nije bilo kada ih je dodirnula nezagrijana sonda.[43] Iako je kod insekata nocicepcija dokazana, ne postoji konsenzus da insekti svjesno osjećaju bol.[44]
Probavni sistem
[uredi | uredi izvor]Insekt koristi svoj probavni sistem za svefaze u preradi hrane: probavu, apsorpciju i isporuku i eliminaciju izmeta.[45][46] Većina hrane unosi se u obliku makromolekula i drugih složenih supstanci poput proteina, polisaharida, masti i nukleinskih kiselina . Te makromolekule moraju se razgraditi pomoću kataboličkih reakcija na manje molekule poput aminokiselina i jednostavnih šećera, prije nego što ih ćelije tijela koriste za energiju, rast ili reprodukciju . Ovaj proces raspadanja poznat je kao probava. Glavna struktura probavnog sistema insekata je dugačka zatvorena cijev zvana probavni kanal (ili crijeva), koja prolazi duž tijela. Probavni kanal usmjerava hranu u jednom smjeru: od usta do anusa. U crijevima se odvija gotovo sva probava insekata. Može se podijeliti u tri dijela - prednje crijevo, srednje crijevo i stražnje crijevo – od kojih svaki vrši drugačiji proces probave.[47] Osim probavnog kanala, insekti imaju i uparene pljuvačne žlijezde i pljuvačne rezervoare. Ove strukture obično se nalaze u toraksu, uz prednje crijevo.[1]
Prednje crijevo
[uredi | uredi izvor]
Prvi odjeljak probavnog kanala je prednje crijevo (element 27 u numeriranom dijagramu), ili stomodaeum. Prednje crijevo je obloženo kutikulskom oblogom od hitina i proteina kao zaštita od žilave hrane. Uključuje usnu šupljinu (usta), ždrijelo, jednjak i voljku i proventrikulus (bilo koji dio može biti visoko modificiran ), koji istovremeno čuvaju hranu i i znak kada treba nastaviti dalje prema srednjem crijevu.[1] Ovdje probava započinje kad se djelomično prožvakana hrana razgrađuje pljuvačkom iz pljuvačnih žlijezda. Dok pljuvačne žlijezde proizvode tečnost i enzim za varenje ugljikohidrata (uglavnom amilaze), snažni mišići u ždrijelu pumpaju tekućinu u usnu šupljinu, podmazujući hranu poput salivarija i pomažući hranilice za krv i hranilice za razgradnju ksilema i floema. Odatle ždrijelo prenosi hranu do jednjaka, koja bi mogla biti obična cijev koja je prenosi u voljku i proventrikulus, a zatim na srednjeg crijeva, kao kod većine insekata. Alternativno, prednja crijeva mogu se proširiti u vrlo povećani voljku i proventrikulus, ili voljka može biti samo divertikulum ili struktura ispunjena tečnošću, kao kod nekih vrsta Diptera.[48]

Pljuvačna žlijezda (element 30 na numeriranom dijagramu) u ustima insekata proizvodi pljuvačku . Pljuvačni kanali vode od žlijezda do rezervoara, a zatim kroz glavu prema otvoru zvanom salivarij, smještenom iza hipofarinksa. Pomičući dijelove usta (element 32 u numeriranom dijagramu) insekt može miješati hranu sa pljuvačkom. Smjesa pljuvačke i hrane zatim putuje kroz pljuvačne cijevi u usta, gdje se počinje razgrađivati.[45][49] Neki insekti, poput muha, imaju vanusnu probava. Insekti koji koriste takvuu probavu izbacuju probavne enzime na hranu kako bi je razgradili. Ova strategija omogućava insektima da iz izvora hrane izvuku značajan dio dostupnih hranjivih sastojaka.[48]:31
Srednje crijevo
[uredi | uredi izvor]Kada hrana napusti voljku, prelazi u srednje crijevo (element 13 na numeriranom dijagramu), poznat i kao mezenteron, u kojem se odvija većina probave. Mikroskopske izbočine iz zida srednjeg crijeva, nazvane mikrovile, povećavaju površinu zida i omogućavaju apsorpciju više hranljivih sastojaka; imaju tendenciju da budu blizu porekla srednjeg crijeva. U nekim insektima uloga mikrovila i gdje se nalaze mogu varirati. Naprimjer, specijalizirane mikrovile koje proizvode probavne enzime mogu vjerovatnije biti blizu kraja srednjeg crijeva, a apsorpcija blizu početka ili na samom početku srednjeg crijeva.[48]
Stražnje crijevo
[uredi | uredi izvor]U zadnjem crijevu (element 16 na numeriranom dijagramu), ili proktodejumu, neprobavljenim česticama hrane pridružuje se mokraćna kiselina kako bi se stvorile izmetne kuglice. Rektum apsorbira 90% vode u tim fekalnim kuglicama, a suha peleta se zatim eliminira kroz anus (element 17), dovršavajući proces probave. Mokraćna kiselina nastaje upotrebom hemolimfnih otpadnih proizvoda difundiranih iz Malpighijevih cjevčica (element 20). Zatim se prazni direktno u probavni kanal, na spoju između srednjeg i stražnjeg crijeva. Broj malpighijevih tubula koje posjeduje određeni insekt varira između vrsta, krećući se od samo dvije kod nekih insekata do preko 100 tubula kod drugih.[1]:71–72, 78–80
Respiratorni sistemi
[uredi | uredi izvor]Disanje insekata odvija se bez pluća. Umjesto toga, insekati imaju sistem unutrašnjih cijevi i vrećica kroz koje se plinovi ili difundiraju ili se aktivno pumpaju, isporučujući kiseik izravno tkivima kojima je potreban putem traheja (element 8 u numeriranom dijagramu). Budući da se kisik isporučuje izravno, cirkulacijski sistem se ne koristi za prijenos kisika, pa je stoga znatno smanjen. Cirkulacijski sistem insekata nema vena ili arterija, a umjesto toga sastoji se od nešto više od jedne, perforirane leđne cijevi koja pulsira peristaltički. Prema toraksu, leđna cijev (element 14) dijeli se na komore i djeluje poput srca insekta. Suprotni kraj leđne cijevi je poput aorte insekta koji cirkulira hemolimfu, analog tečnosti člankonožaca krvi, unutar tjelesne šupljine.[1]:61–65[50] Zrak se uvlači kroz otvore na bočnim stranama trbuha zvane spirakule.
Postoje mnoge različite sheme razmjene plinova koje pokazuju različite grupe insekata. Uzorci razmene gasova kod insekata mogu se kretati od kontinuirane i difuzne ventilacije do diskontinuirane razmene plinova.[1] Tokom kontinuirane razmene plinova, kisik se unosi, a ugljik-dioksid oslobađa u kontinuiranom ciklusu. Međutim, u prekidnoj razmjeni plinova, insekt uzima kisik dok je aktivan, a male količine ugljik-dioksida oslobađaju se dok miruje.[51] Difuzna ventilacija je jednostavno oblik kontinuirane izmjene plina koja se događa difuzijom, a ne fizičkim unosom kisika. Neke vrste insekata koji su potopljeni takođe imaju prilagodbe koje pomažu u disanju. Kao larve, mnogi insekti imaju škrge koje mogu izdvojiti kiseonik rastvoren u vodi, dok se drugi trebaju podići na površinu vode kako bi nadoknadili zalihe zraka, koje mogu biti zadržane ili zarobljene u posebnim strukturama.[52][53]
Cirkulacijski sistem
[uredi | uredi izvor]Tjelesna tečnost („krv“) insekata ili hemolimfa glavna je funkcija transporta i oplakuje tjelesne organe insekata. Sačinjavajući obično manje od 25% tjelesne težine insekta, ona prenosi hormone, hranjive sastojke i otpad i ima ulogu u osmoregulaciji, kontroli temperature, imunosti, skladištenju (voda, ugljikohidrati i masti) i skeletna funkcija. Takođe ima bitnu ulogu u procesu presvlačenja.[54][55] Pored toga, hemolimfa un nekim redovima može služiti za odbranu od neprtijatelja. Sadrži neukusne i neprijatne hemikalije koje će djelovati kao odvraćanje predatora.[1] Hemolimfa sadrži molekule, ione i ćelije;[1] regulira razmjenu hemikalija između tkiva. Zatvorena je u tjelesnoj šupljini insekta ili hemocelu.[1][56] Prenosi se po tijelu kombiniranim srčanim (stražnjim) i aortnim (prednjim) pulsacijama, koje se nalaze leđno tik ispod površine tijela.[1][54][55] Razlikuje se od kičmenjačke krvi po tome što ne sadrži crvene krvne ćelije i samim tim nema visoku nosivost kisika, a sličnija je limfi kičmenjaka.[1][56]
Tjelesne tečnosti ulaze kroz jednosmjerne zaliske ostije, a to su otvori smješteni dužinom kombinirane aorte i srčanog organa. Do pumpanja hemolimfe dolazi talasima peristaltičke kontrakcije, sa stražnjeg kraja tijela, pumpajući unazad u leđni sud, van aorte, a zatim u glavu gdje odlazi u hemocel.[1]< ref name = "Elzinga, 2004"/> Hemolimfa cirkulira u dodatke jednosmjerno uz pomoć mišićnih pumpi ili pomoćnih pulsirajućih organa koji se obično nalaze u osnovi antene ili u krilima, a ponekad i u nogama,[1] sa brzinom pumpanja koja se ubrzava s periodima povećane aktivnosti.[55] Kretanje hemolimfe posebno je važno za termoregulaciju u redovima kao što su Odonata , Lepidoptera, Hymenoptera i Diptera.[1]
Endokrini sistem
[uredi | uredi izvor]Slijedeće žlijezde su dio endokrinog sistema:
1. Neurosekretorne ćelije 2. Corpora cardiaca 3. Protoraksna žlijezda 4. Corpora allata[57][58]
Reproduktivni sistem
[uredi | uredi izvor]Ženski
[uredi | uredi izvor]Ženski insekti mogu stvarati jaja, primati i skladištiti spermu, manipulirati spermom različitih mužjaka i odlagati jaja. Njihov reproduktivni sistem sastoji se od para jajnika, pomoćnih žlijezda, jedne ili više spermateka i kanala koji povezuju ove dijelove. Jajnici stvaraju jaja, a pomoćne žlijezde proizvode supstance koje pomažu u pakiranju i polaganju jaja. Spermateke čuvaju spermu u različitim vremenskim periodima i, zajedno sa dijelovima jajovoda, mogu kontrolirati upotrebu sperme. Kanali i spermateke obloženi su kutikulom.[8]:880
Jajnici se sastoje od određenog broja jajnih cijevi, nazvanih ovariole, koje se razlikuju u veličini i broju prema vrstama. Broj jaja koje insekt može stvoriti varira u zavisnosti od broja ovariola, s tim da na brzinu razvoja jaja utiče i dizajn ovariola. U meroističnim jajnicima, buduća jaja se uzastopno dijele i većina ćelija ćerki postaju pomoćne ćelije za jedan oocit u klasteru. U panoističkim jajnicima, svako jaje koje će proizvesti matične ćelijske klice razvija se u oocit; nema pomoćnih ćelija s klicine linije.
Pomoćne žlijezde ili žljezdani dijelovi jajovoda proizvode razne supstance za održavanje, transport i oplodnju sperme, kao i za zaštitu jajnih ćelijaa. Mogu proizvesti ljepilo i zaštitne supstance za oblaganje jaja ili čvrste obloge za seriju jaja koja se nazivaju ooteke. Spermateke su cijevi ili vrećice u kojima se sperma može čuvati između vremena parenja i trenutka oplodnje jajne ćelije. Ispitivanje očinstva insekata otkrilo je da neki, a vjerovatno i mnogi ženski insekti koriste spermateku i razne kanale za kontrolu ili sprečavanje sperme koja se koristi u korist nekih mužjaka u odnosu na druge.[8]
Mužjak
[uredi | uredi izvor]Glavna komponenta muškog reproduktivnog sistema je testis, podržan u tjelesnoj šupljini pomoću trahejama i masnim tijelom. Primitivniji insekti bez krila imaju jedan testis, a u nekih Lepidoptera dva testisa koji sazrijevaju sekundarno su stopljeni u jednu strukturu tokom kasnijih faza razvoja larvi, iako kanali koji vode iz njih ostaju odvojeno. Međutim, većina muških insekata ima par testisa, unutar kojih su sjemenici ili folikule zatvoreni u opnastu vrećicu. Folikule se pomoću vas eferens povezuju s vas deferens, a dvije cjevaaste vas deferens povezuju se sa srednjim ejakulacijskim kanalom, koji vodi prema van. Dio sjemenovoda često je uvećan da bi formirao sjemensku vezikulu, koja čuva spermu prije nego što se ispusti u ženku. Sjemene vezikule imaju žljezdane obloge koje izlučuju hranjive sastojke za prehranu i održavanje sperme. Ejakulacijski kanal izveden je iz invaginacije ćelija epiderme tokom razvoja i, kao rezultat toga, ima kutikulsku sluznicu. Krajnji dio ejakulacijskog kanala može se sklerotizirati da bi se stvorio intromitentni organ, edeagus. Ostatak muškog reproduktivnog sistema izveden je iz embrionskog mezoderm, osim zametnih ćelija ili spermatogonija, koje se vrlo rano spuštaju iz matičnih polarnih ćelija tokom embriogeneze.[8] Edeagus može biti prilično izražen ili de minimis. Njegova aza može biti djelimično sklerotizirana faloteka, koja se naziva i falosoma ili teka. Kod nekih vrsta faloteka sadrži prostor, nazvan endosom (unutrašnja vrećica za držanje), u koji se vrh edeagusa može povući. Vas deferens se ponekad uvlači (presavijen) u faloteku zajedno sa sjemenskom vezikulom.[59][60]
Također pogledajte
[uredi | uredi izvor]- Morfologija (biologija)
- Fiziologija insekata
- Ekologija insekata
- Let insekata
- Beskičmenjaci
- Insekti
- Entomologija
- Prahistorijski insekti
Reference
[uredi | uredi izvor]- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag Gullan, P.J.; P.S. Cranston (2005). The Insects: An Outline of Entomology (3 izd.). Oxford: Blackwell Publishing. ISBN 1-4051-1113-5.
- ^ "O. Orkin Insect zoo". The University of Nebraska Department of Entomology. Arhivirano s originala, 2. 6. 2009. Pristupljeno 3. 5. 2009.
- ^ Resh, Vincent H.; Cardé, Ring T. (2009). Encyclopedia of Insects (2nd izd.). San DIego, CA: Academic Press. str. 12.
- ^ Campbell, N. A. (1996) Biology (4th edition) Benjamin Cummings, New Work. p. 69 ISBN 0-8053-1957-3
- ^ Gene Kritsky. (2002). A Survey of Entomology. iUniverse. ISBN 978-0-595-22143-1.
- ^ "external morphology of Insects" (PDF). Arhivirano s originala (PDF), 19. 7. 2011. Pristupljeno 20. 3. 2011.
- ^ a b Richards, O. W.; Davies, R.G. (1977). Imms' General Textbook of Entomology: Volume 1: Structure, Physiology and Development Volume 2: Classification and Biology. Berlin: Springer. ISBN 0-412-61390-5.
- ^ a b c d e f g h i j k l m Resh, Vincent H.; Ring T. Carde (2009). Encyclopedia of Insects (2 ed.). U. S. A. Academic Press. ISBN 978-0-12-374144-8.CS1 održavanje: upotreba parametra authors (link)
- ^ Smith, John Bernhard, Explanation of terms used in entomology Publisher: Brooklyn entomological society 1906 (May be downloaded from: https://archive.org/details/explanationofter00smit)
- ^ Fox, Richard (6 Oct 2006). "External Anatomy". Lander University. Arhivirano s originala, 14. 3. 2011. Pristupljeno 20. 3. 2011.
- ^ "Archived copy" (PDF). Arhivirano s originala (PDF), 1. 10. 2008. Pristupljeno 20. 3. 2011. journal zahtijeva
|journal=(pomoć)CS1 održavanje: arhivirana kopija u naslovu (link) - ^ Meyer-Rochow, V.B. (1974). "Structure and function of the larval eye of the sawfly larva Perga". Journal of Insect Physiology. 20 (8): 1565–1591. doi:10.1016/0022-1910(74)90087-0. PMID 4854430.
- ^ Völkel, R.; Eisner, M.; Weible, K.J. (2003). "Miniaturized imaging systems". Microelectronic Engineering. 67–68: 461–472. doi:10.1016/S0167-9317(03)00102-3. ISSN 0167-9317.
- ^ Gaten, Edward (1998). "Optics and phylogeny: is there an insight? The evolution of superposition eyes in the Decapoda (Crustacea)". Contributions to Zoology. 67 (4): 223–236. doi:10.1163/18759866-06704001.
- ^ Ritchie, Alexander (1985). "Ainiktozoon loganense Scourfield, a protochordate? from the Silurian of Scotland". Alcheringa. 9 (2): 137. doi:10.1080/03115518508618961.
- ^ Mayer, G. (2006). "Structure and development of onychophoran eyes: What is the ancestral visual organ in arthropods?". Arthropod Structure and Development. 35 (4): 231–245. doi:10.1016/j.asd.2006.06.003. PMID 18089073.
- ^ a b c d e Chapman, R.F. (1998). The Insects: Structure and Function (4th izd.). Cambridge, UK: Cambridge University Press. ISBN 0521570484.
- ^ Krause, A.F.; Winkler, A.; Dürr, V. (2013). "Central drive and proprioceptive control of antennal movements in the walking stick insect". Journal of Physiology, Paris. 107 (1–2): 116–129. doi:10.1016/j.jphysparis.2012.06.001. PMID 22728470.
- ^ Okada, J; Toh, Y (2001). "Peripheral representation of antennal orientation by the scapal hair plate of the cockroach Periplaneta americana". Journal of Experimental Biology. 204 (Pt 24): 4301–4309. PMID 11815654.
- ^ Staudacher, E.; Gebhardt, M.J.; Dürr, V. (2005). "Antennal movements and mechanoreception: Neurobiology of active tactile sensors". Advances in Insect Physiology. 32: 49–205. doi:10.1016/S0065-2806(05)32002-9. ISBN 9780120242320.
- ^ Servadei, A.; Zangheri, S.; Masutti, L. (1972). Entomologia generale ed applicata. CEDAM. str. 492–530.
- ^ Campbell, Frank L.; Priestly, June D. (1970). "Flagellar Annuli of Blattella germanica (Dictyoptera: Blattellidae).–Changes in Their Numbers and Dimensions during Postembryonic Development". Annals of the Entomological Society of America. 63: 81–88. doi:10.1093/aesa/63.1.81.
- ^ "Insect antennae". The Amateur Entomologists' Society. Pristupljeno 21. 3. 2011.
- ^ "Insect Morphology". University of Minisotta (Department of Entomology). Arhivirano s originala, 3. 3. 2011. Pristupljeno 21. 3. 2011.
- ^ Kirejtshuk, A.G. (novembar 2002). "Head". Beetles (Coleoptera) and coleopterologist. zin.ru. Pristupljeno 21. 3. 2011.
- ^ a b c d e f g Chapman, R.F. (1998). The Insects: Structure and function (4th izd.). Cambridge, New York: Cambridge University Press. ISBN 0-521-57048-4.
- ^ a b c Gilliott, Cedric (august 1995). Entomology (2 izd.). Springer-Verlag New York, LLC. ISBN 0-306-44967-6.
- ^ Kapoor, V.C. C. (januar 1998). Principles and Practices of Animal Taxonomy. 1 (1 izd.). Science Publishers. str. 48. ISBN 1-57808-024-X.
- ^ "Mosquito biting mouthparts". allmosquitos.com. 2011. Pristupljeno 17. 4. 2011.
- ^ a b c Scoble, MJ. (1992). The Lepidoptera: Form, function, and diversity. Oxford Univ. Press. ISBN 978-1-4020-6242-1.
- ^ Heppner, J. B. (2008). "Butterflies and moths". u Capinera, John L. (ured.). Encyclopedia of Entomology. Gale virtual reference library. 4 (2nd izd.). Springer Reference. str. 4345. ISBN 978-1-4020-6242-1. Upotreblja se zastarjeli parametar
|chapterurl=(pomoć) - ^ Agosta, Salvatore J.; Janzen, Daniel H. (2004). "Body size distributions of large Costa Rican dry forest moths and the underlying relationship between plant and pollinator morphology". Oikos. 108 (1): 183–193. doi:10.1111/j.0030-1299.2005.13504.x.
- ^ Kunte, Krushnamegh (2007). "Allometry and functional constraints on proboscis lengths in butterflies" (PDF). Functional Ecology. 21 (5): 982–987. doi:10.1111/j.1365-2435.2007.01299.x. Arhivirano s originala (PDF), 29. 6. 2011. Pristupljeno 26. 2. 2011.
- ^ "Sponging". University of Minissota. Arhivirano s originala, 27. 9. 2011. Pristupljeno 17. 4. 2011.
- ^ "Fly Mouthparts". School of Biological Sciences Online Learning Resources. University of Sydney. 4. 2. 2010. Arhivirano s originala, 9. 4. 2011. Pristupljeno 17. 4. 2011.
- ^ Meyer, John R. (5. 1. 2007). "External Anatomy: WINGS". Department of Entomology, NC State University. Arhivirano s originala, 16. 7. 2011. Pristupljeno 21. 3. 2011.
- ^ a b c d e f g h i j k l m n o p q r Snodgrass, R. E. (decembar 1993). Principles of Insect Morphology. Cornell Univ Press. ISBN 0-8014-8125-2.
- ^ Spieth, HT (1932). "A New Method of Studying the Wing Veins of the Mayflies and Some Results Therefrom (Ephemerida)" (PDF). Entomological News. Arhivirano s originala (PDF), 30. 9. 2011.
- ^ "profemur, profemora - BugGuide.Net". bugguide.net. Pristupljeno 10. 12. 2016.
- ^ "protibia - BugGuide.Net". bugguide.net. Pristupljeno 10. 12. 2016.
- ^ Schneiderman, Howard A. (1960). "Discontinuous respiration in insects: role of the spiracles". Biol. Bull. 119 (3): 494–528. doi:10.2307/1539265. JSTOR 1539265.
- ^ Eisemann, WK; Jorgensen, W. K.; Merritt, D. J.; Rice, M. J.; Cribb, B. W.; Webb, P. D.; Zalucki, M. P.; et al. (1984). "Do insects feel pain? — A biological view". Cellular and Molecular Life Sciences. 40 (2): 1420–1423. doi:10.1007/BF01963580.
- ^ Tracey, J; Wilson, RI; Laurent, G; Benzer, S; et al. (18. 4. 2003). "painless, a Drosophila gene essential for nociception". Cell. 113 (2): 261–273. doi:10.1016/S0092-8674(03)00272-1. PMID 12705873.
- ^ Sømme, LS (14. 1. 2005). "Sentience and pain in invertebrates". Norwegian Scientific Committee for Food Safety. Pristupljeno 30. 9. 2009.[trajno mrtav link]
- ^ a b "General Entomology - Digestive and Excritory system". NC state University. Pristupljeno 3. 5. 2009.
- ^ Terra, Walter R.; Ferreira, Clélia (2009), "Digestive System", Encyclopedia of Insects (jezik: engleski), Elsevier, str. 273–281, doi:10.1016/b978-0-12-374144-8.00083-7, ISBN 978-0-12-374144-8, pristupljeno 29. 9. 2020
- ^ Terra, Walter R.; Ferreira, Clélia (1. 1. 2009), "Chapter 74 - Digestive System", u Resh, Vincent H.; Cardé, Ring T. (ured.), Encyclopedia of Insects (Second Edition) (jezik: engleski), San Diego: Academic Press, str. 273–281, doi:10.1016/b978-0-12-374144-8.00083-7, ISBN 978-0-12-374144-8, pristupljeno 29. 9. 2020
- ^ a b c Nation, James L. (novembar 2001). "15". Insect Physiology and Biochemistry (1 izd.). CRC Press. str. 496pp. ISBN 0-8493-1181-0.
- ^ Duncan, Carl D. (1939). A Contribution to The Biology of North American Vespine Wasps (1 izd.). Stanford: Stanford University Press. str. 24–29.
- ^ Meyer, John R. (17. 2. 2006). "Circulatory System". NC State University: Department of Entomology, NC State University. str. 1. Arhivirano s originala, 27. 9. 2009. Pristupljeno 11. 10. 2009.
- ^ Chown, S.L.; S.W. Nicholson (2004). Insect Physiological Ecology. New York: Oxford University Press. ISBN 0-19-851549-9.
- ^ Richard W. Merritt, Kenneth W. Cummins, and Martin B. Berg (editors) (2007). An Introduction to the Aquatic Insects of North America (4th izd.). Kendall Hunt Publishers. ISBN 978-0-7575-5049-2.CS1 održavanje: više imena: authors list (link) CS1 održavanje: dodatni tekst: authors list (link)
- ^ Merritt, RW, KW Cummins, and MB Berg (2007). An Introduction To The Aquatic Insects Of North America. Kendall Hunt Publishing Company. ISBN 978-0-7575-4128-5.CS1 održavanje: više imena: authors list (link)
- ^ a b McGavin, G. C. (2001). Essential Entomology; An order by order introduction. New York: Oxford University Press. ISBN 9780198500025.
- ^ a b c Triplehorn, C. A.; Johnson, N. F. (2005). Borror and DeLong's Introduction to the Study of Insects. Brooks / Thomson Cole: Brooks / Thomson Cole.
- ^ a b Elzinga, R.J. (2004). Fundamentals of Entomology (6th izd.). New Jersey USA: Pearson/Prentice Hall.
- ^ Triplehorn, Charles A; Johnson, Norman F (2005). Borror and DeLong's introduction to the study of insects (7th izd.). Australia: Thomson, Brooks/Cole. ISBN 9780030968358.
- ^ Gullan, P.J.; P.S. Cranston (2005). The Insects: An Outline of Entomology (3 izd.). Oxford: Blackwell Publishing. str. 61–65. ISBN 1-4051-1113-5.
- ^ De Carlo; J. A. (1983). "Hemipteros acuáticos y semiacuáticos. Estudio en grupos en las partes de igual función de los aparatos genitales masculinos de especies estudiadas". Revista de la Sociedad Entomológica Argentina. 42 (1/4): 149–154.
- ^ Andersen, N. Møller (1991). "Marine insects: genital morphology, phylogeny and evolution of sea skaters, genus Halobates (Hemiptera: Gerridae)". Zoological Journal of the Linnean Society. 103 (1): 21–60. doi:10.1111/j.1096-3642.1991.tb00896.x.
Vanjski linkovi
[uredi | uredi izvor]| Sisari |  | |
|---|---|---|
| Ostali kičmenjaci | ||
| Beskičmenjaci | ||
| Biljke | ||
| Discipline |
| |
| Ostale teme | ||