Mesajcı RNA
Mesajcı RNA (İngilizce: messenger RNA) (mRNA), sentezlenecek bir proteinin amino asit dizisine karşılık gelen kimyasal şifreyi taşıyan bir moleküldür. mRNA, bir DNA kalıptan transkripsiyon yoluyla sentezlenir ve protein sentez yeri olan ribozomlara, protein kodlayıcı bilgiyi taşır. Burada, çevirim (translasyon) süreci sonucu, RNA polimerindeki bilgi ile bir amino asit polimeri üretilir. Nükleik asitlerin (mRNA ve DNA) amino asit dizilerine karşılık gelen bölgelerindeki her üç baz, proteindeki bir amino asite karşılık gelir. Bu üçlülere kodon denir, her biri bir amino asit kodlar, bitiş kodonu ise protein sentezini durdurur. Bu işlem iki diğer RNA türünü daha gerektirir: taşıyıcı RNA (tRNA) kodonun tanınmasına aracılık eder ve ona karşılık gelen amino asiti getirir; ribozomal RNA (rRNA) ise ribozomdaki protein imalat mekanizmasının kataliz merkezidir.
Sentez, işlem ve fonksiyon
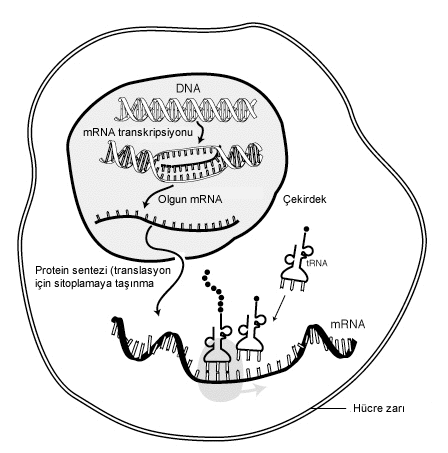
değiştirBir mRNA molekülünün varlığı onun transkripsiyonu ile başlar ve yıkımı ile sona erer. Çevrim başlamadan evvel bir mRNA molekülü işlenmiş, redaksiyona uğramış ve taşınmış olabilir. Ökaryotik mRNA molekülleri çoğu zaman kapsamlı bir işlenme (İng. procesing) ve taşınma gerektirirler, prokaryotlar ise bunu gerektirmezler.
Transkripsiyon
değiştirTranskripsiyon sırasında RNA polimeraz, DNA'daki bir gendeki bilgiyi gereken sayıda mRNA olarak kopyalar. Bu süreç ökaryot ve prokaryotlarda birbirine benzer. Önemli bir fark, transkripsiyon sırasında ökaryotik RNA polimerazın mRNA işlenme enzimleriyle beraber olmasıdır, öyle ki transkripsiyon başladıktan sonra işlenme hızla takip edebilir. Kısa ömürlü olan, işlenmemiş veya kısmen işlenmiş ürüne pre-mRNA denir; tamamen işlendikten sonra olgun mRNA olarak adlandırılır.
Ökaryotik pre-mRNA işlemesi
değiştirmRNA işlemesi ökaryot, prokaryot ve arkeler arasında büyük farklılık gösterir. Ökaryotik olmayan mRNA'lar transkripsiyon sonrasında olgundurlar ve ender durumlar dışında işlenme gerektirmezler. Buna karşın, ökaryotik pre-mRNA'lar, kapsamlı bir işlenme gerektirirler.
5' başlık eklemesi
değiştir5' başlık (diğer adlarıyla RNA başlığı, RNA 7-metilguanozin başlığı veya RNA m7G başlık), transkripsiyon başlangıcından hemen sonra bir ökaryotik mRNA'nın "önüne" yani 5' ucuna eklenmiş, değişime uğramış bir guanin bazlı bir nükleotittir. 5' başlık, ilk yazılmış nükleotit ile 5'-5'-trifosfat bağı ile bağlanmış bir uç (terminal) 7-metilguanozin kalıntısıdır. mRNA'nın ribozom tarafından tanınması ve RNaz'lardan korunması için varlığı çok önemlidir.
Başlık eklemesi transkripsiyon eşliklidir ve transkripsiyonla beraber gerçekleşir, öyle ki iki süreç birbirlerine etki ederler. Transkripsiyonun başlamasından kısa süre sonra, sentezlenmekte olan mRNA'nın 5' ucuna, RNA polimeraz ile birleşik olan başlık sentezleme kompleksi bağlanır. Bu enzim kompleksi, mRNA başlık eklemesi için gerekli kimyasal adımları katalizler. Sentez, çok adımlı bir tepkimedir.
Uçbirleştirme
değiştirUçbirleştirme (İng. splicing), intron adı verilen, protein kodlamayan bazı kısımların pre-mRNA'dan çıkartılması işlemidir; kalan kısımlar protein kodlayıcı dizileri kapsar ve bunlara ekson denir. Bazen pre-mRNA molekülleri farklı şekillerde uçbirleştirmeye uğrayabilirler, böylece bir gen birden fazla proteini kodlayabilir. Bu sürece alternatif uçbirleştirme denir. Uçbirleştirme genelde splisozom olarak adlandırılan bir RNA-protein kompleksi tarafından gerçekleştirir, ama bazı RNA molekülleri kendi uçbirleştirmelerini kendileri katalizleyebilir. (bkz. ribozimler).
Redaksiyon
değiştirBazı durumlarda bir mRNA'nın bilgi içeriği, onu meydana getiren bazlarda kimyasal değiklikler sonucu, hücre tarafından değiştirilir. İnsanlarda bunun bir örneği, apolipoprotein B mRNA'sıdır, bazı dokularda dizisi değişir (redakte olur), bazılarında ise değişmez. Redaksiyon sonucu erken bir durma kodonu yaratılır, bu da çevrim sonucu daha kısa bir protein oluşmasına neden olur.
Poliadenilasyon
değiştirPoliadenilasyon, bir mRNA molekülüne bir poladenilil parçasının kovalent bağ ile eklenmesidir. Ökaryotik organizmalarda çoğu mRNA, 3' ucunda poliadenile olur. Poli(A) kuyruğu ve ona bağlı olan protein, mRNA'nın eksonükleazlar tarafından yıkımından korur. Poliadenilasyon ayrıca, transkripsiyon sonlanması için, mRNA'nın çekirdekten ihracı ve çevirim için de önemlidir. mRNA prokaryotik organizmlarda da poliadenile olabilir, ama bu canlılarda poli(A) kuyruğu eksonükleotik yıkımı engellemek yerine kolaylaştırır.
Poliadenilasyon DNA'nın RNA'ya yazılımı sırasında ve onu hemen takiben olur. Transkripsiyon tamamlanmasının ardından RNA polimerazla birleşik bir endonükleaz kompleksi aracılığıyla mRNA zinciri kesilir. mRNA kesildikten sonra kesim yerindeki serbest 3' uca yaklaşık 250 adenozin kalıntısı eklenir. Alternatif uçbirleştirmede olduğu gibi, bir mRNA'nın birden fazla sayıda poliadenilasyon çeşidi olabilir.
Taşıma
değiştirÖkaryotlarla prokaryotlar arasındaki bir diğer fark mRNA taşımasıdır. Ökaryotik transkripsiyon ve translasyon hücre içinde farklı bölmelerde cereyan ettiği için, ökaryotik mRNA'nın hücre çekirdeğinden sitoplazmaya ihracı gerekmektedir. Olgun mRNA'lar gördükleri işlemler sayesinde tanınırlar ve çekirdek gözeneğinden ihraç edilirler. Nöronlarda mRNA'lar somadan dendritlere taşınırlar, dış bir uyarmanın ardından orada yerel bir çevirim olur.[1] Çoğu mesaj ise kendilerini hücre içinde belli konuma hedefleyen "posta kodları" taşır.[2]
Çevirim
değiştirProkaryotlarda mRNA işlenme ve taşınmaya gerek duymadığı için ribozom tarafından çevrilmeleri transkripsiyon olur olmaz başlayabilir. Bu yüzden prokaryotik çevrimin transkripsiyonla eşlikli ve "ko-transkripsiyonel" (eş yazımlı) olduğu söylenir.
İşlenmiş ve sitoplazmaya taşınmış ökaryotik mRNA (yani olgun mRNA) ribozomlar tarafından çevrime uğrayabilir. Çevrim ya sitoplazmada serbest dolaşan ribozomlarda olur ya da, sinyal tanıma taneciği mRNA'yı endoplazmik retikuluma havale ettikten sonra oradaki ribozomlarda olur. Dolayısıyla, prokaryotlardan farklı olarak, ökaryotik çevrim, transkripsiyon ile doğrudan eşlikli değildir.
Yapı
değiştir5' başlık
değiştir5' başlık pre-mRNA'nın "önüne" (5' ucuna) 5'-5'-trifosfat bağı ile eklenmiş, değişime uğramış bir guanin nükleotittir. Bu değişim mRNA'nın ribozomar tarafından tanınıp bağlanması, ayrıca 5' eksonükleazlardan korunması için gereklidir. Uçbirleştirme ve taşınma gibi başka temel süreçler için de gereklidir.
Kodlayıcı bölgeler
değiştirKodlayıcı bölgeler kodonlardan oluşur, bunlarda bulunan bilgi deşifre edilip ribozomlar tarafından bir proteine (genelde ökaryotlarda) veya birkaç proteine (genelde prokaryotlarda) çevrilir. Kodlayıcı bölgeler bir başlama kodonu ile başlar ve bir bitiş kodonu ile sona erer. Genelde başlama kodonu bir AUG üçlüsüdür, bitiş kodonu ise UAA, UAG veya UGA'dır. Kodlayıcı bölge genelde dahilî baz çiftleriyle stabilize durumdadır, bu onun yıkımını engeller.[3][4] Protein kodlayıcı olmanın yanı sıra, kodlayıcı bölgenin bazı kısımları pre-mRNA'dadüznleyici dizi olarak işlev görebilirler, örneğin eksonik uçbirlştirme hızlandırıcısı ve eksonik uçbirleşirme susturucusu olarak.
Çevrilmeyen bölgeler
değiştirÇevrilmeyen bölgeler (yaygın olarak İngilizce Untranslated region kısaltması olan UTR olarak adlandırılırlar) mRNA'nın başlama kodonundan evvel ve bitiş kodonundan sonra gelen, çevrilmeyen kısımlardır. Bunlar kodlayıcı bölge ile birlikte yazılırlar, olgun RNA'da da yer aldıkları için eksonik sayılırlar. Bu bölgelerin gen ifadesinde çeşitli roller oynadıkları gösterilmiştir, bunların arasında mRNA kararlılığı (stabilitesi), mRNA konumlanması (lokalizasyonu) ve çevirim verimliliği bulunmaktadır. Bir UTR'nin bu işlevleri yerine getirmesi UTR'de bulunan dizilere bağlıdır ve mRNA'lar arasında fark edebilir.
mRNA'nın kararlılığının 5' UTR ve/veya 3' UTR tarafından belirlenebilmesinin nedeni, ribonükleaz denen RNA yıkıcı enzimlerin ve yardımcı proteinlerinin onlara farklı bağlanma afiniteleri göstermesidir.
Çevirim verimliliği ve bazen çevirimin tamamen ketlenmesi (inhibisyonu) UTR'ler tarafından kontrol edilebilir. 3' veya 5' UTR'lere bağlanan proteinler ribozomların mRNA'ya bağlanmasına etki edebilirler. 3' UTR'ye bağlanan mikroRNA'lar da çevirim verimliliğini ve mRNA kararlılığını etkileyebilir.
mRNA'nın sitoplazmada konumlanmasının 3' UTR'nin bir işlevi olduğu düşünülmektedir. Hücrenin belli bir bölgesinde gerekli olan proteinler orada sentezlenebilir; böyle durumlarda 3' UTR, mRNA'nın çevrimi için oraya taşınmasını sağlayacak diziler içerebilir.
Çevrilmeyen bölgelerdeki dizi elemanlarının bazıları RNA'ya yazıldıkları zaman karakteristik ikincil yapılar oluştururlar. Bu yapısal mRNA elemanlar RNA'nın düzenlenmesine yararlar. mRNA elemanlarının bir sınıfı, riboanahtarlar, küçük moleküllere doğrudan bağlanıp şekillerini değiştirirler, bunun sonucu yazılım veya çevirim hızı değişir. Böyle durumlarda, düzenleyici proteinler olaya karışmadığı için, mRNA kendi kendini düzenlemiş olur.
Poli(A) kuyruğu
değiştir3' poli(A) kuyruğu, pre-mRNA'nın 3' kuyruğunda yer alan, çoğu zaman birkaç yüz adenin nükleotidinden oluşmuş bir zincirdir. Bu kuyruk çekirdekten ihracı ve çevirimi sağlar, ayrıca ökaryotlarda mRNA'yı yıkımdan korur.
Monosistronik ve polisistronik mRNA
değiştirBir mRNA tek bir proteini kodlamaya yetecek genetik bilgi içerirse onun monosistronik olduğu söylenir. Çoğu ökaryotik mRNA böyledir.[5]. Buna karşın, polisistronik mRNA'lar birkaç gene ait bilgiyi taşırlar, bunlardan birkaç farklı protein kodlar. Bu proteinler çoğu zaman birbiriyle ilişkili işlevlere sahiptir, bir operon olarak yaynyana bulunurlar ve beraber düzenlenirler. Bakteri ve arkelerde bulunan mRNA'ların çoğu polisistroniktir.[5]. Sadece iki protein kodlayan mRNA için disitronik terimi kullnaılır.
mRNA daireselleşmesi
değiştirÖkaryotlarda başlık bağlayıcı kompleksi ve poli(A)-bağlayıcı proteinin birleşmesi sonucu mRNA'ların daireselleştiği düşünülmektedir.[6] Daireselleşmenin ribozomların geri dönüşümünü sağlayarak çevrimin daha verimli olmasını sağladığı öne sürülmüştür.
Yıkım
değiştirAynı hücrede farklı mRNA'ların farklı ömürleri (stabiliteleri) vardır. Bakterilerde mRNA'ların ömrü birkaç saniyeden bir saatten fazlaya kadar sürebilir; memeli hücrelerinde mRNA ömürleri birkaç dakikadan günlere kadar uzanabilir. Bir mRNA'nın stabilitesi ne kadar yüksek olursa ondan o kadar daha fazla protein üretilebilir. mRNA'nın sınırlı ömrü, hücrenin değişen ihtiyaçlarına bağlı olarak protein sentezini hızla değiştirebilmesini sağlar. Mesajcı RNA'nın ımhasına yol açan çeşitli mekanizmalar vardır, bazıları aşağıda açıklanmıştır.
Prokaryotik mRNA yıkımı
değiştirProkaryotlardaki mRNA'nın ömrü ökaryotlardakinden genelde çok daha kısadır. Prokaryotlarda mRNA ömrünün düzenlenmesi ökaryotlardakinden çok daha basittir. Prokaryotlarda mesajları hızla yıkan çeşitli RNaz'lar vardır, bunlar RNA'nın dizisinden bağımsız olarak etkirler. Alternatif olarak, uzunluğu on ila yüzlerce nükleotit arası olan kısa RNA molekülleri (sRNA) belli mRNA'lara bağlanıp onların yıkımına neden olurlar. sRNA'daki tümleyici (komplemanter) diziler mRNA'ya bağlanarak iki iplikli RNA molekülleri oluştururlar, bunlar bazı tip RNaz'lara substrattırlar.
Yakın zamanda bakteri mRNA'larında da bir çeşit 5' başlık olduğu bulunmuştur. 5' uçta bir trifosfat grubu bulunmaktadır, bundan iki fosfat çıkartılırsa kalan 5' monofosfat mRNA, RNaz E adlı eksonükleaz tarafından parçalanır.[7]
Ökaryotik mRNA devir hızı
değiştirÖkaryotlarda çevirim ile mRNA yıkımı arasında bir denge vardır. Aktif olarak çevrilmekte olan mesajlara polizomlar, ökaryotik başlama faktörleri (eIF-4E ve eIF-4G) ve poli(A) bağlayıcı protein bağlıdır. eIF-4E ve eIF-4G başlık çıkarma enzimi DCP2'yi bloke eder, poli(A) bağlayıcı protein de eksozom kompleksi bloke ederek mesajı korur. Gıda yetersizliği veya viral enfeksiyon durumlarında çevirim aksayabilir ve yıkım hızlanabilir. Çevirim ve yıkım arasındaki denge, P-cismi adı verilen sitoplazmik yapıların büyüklüğü ve çokluğu ile anlaşılabilir.[8] Bir mRNA'nın her çevirimi sırasında poli-A kuyruğu eksonükleazlar tarafından kısaltılır. Bunun sonucu olarak mesajın dairesel yapısı bozulur ve başlık bağlanma yapısı destabilize olur. Mesaj sonra eksozom kompleksi veya başlık çıkartma kompleksi tarafından yıkılır. Böylece etkin olmayan mesajlar hızle yok edilir, aktif mesajlar ise sağlam kalır, hücrenin o anda gerek duyduğu mesajlar seçime uğrar. Çevrimin durması ve mesajın yıkım komplekslerine aktarılmasının mekanizmasının ayrıntıları henüz bilinmemektedir.
AU-zengini eleman yıkımı
değiştirBazı memeli mRNA'larındaki AU-zengini elemanların varlığı, bu dizilere bağlanan proteinlerin etkisi sonucu bu mesajları destabilize eder. AU-zengini elemanlar aracılığıyla mRNA'nın hızlı yıkımı, tümör nekroz faktör ve granülosit-macrofaj koloni uyarıcı faktör (GM-CSF) gibi etkili sitokinlerin fazla üretilmesine engel olan önemli bir mekanizmadır.[9] AU-zengini elemanlar ayrıca c-Jun ve c-Fos gibi onkogenik transkripsiyon faktörlerini düzenler.[10] AU-zengini elemanları tanıyan bağlanıcı proteinler hem eksozom kompleksi[11] hem de başlık çıkarma kompleksi[12] tarafından yıkıma önayak olur.
Anlamsızlık aracılıklı yıkım
değiştirÖkaryot mesajlar, mesaj içinde erken durma kodonlarına (anlamsız kodonlara) bakan "anlamsızlık aracılıklı yıkım" (İng. nonsense mediated decay) ile denetlenirler. Bu durma kodonları alternatif uçbirleştirme, adaptif immün sistemdeki V(D)J rekombiansyonu, DNA'da mutasyonlar, transkripsiyon hataları gibi nedenlerle meydana gelebilir. Erken bir bitiş kodonunun algılanması halinde, ya 5 uçtan başlık çıkarma kompleksi ile, ya 3' uçtan eksozom ile ya da endonükleolitik kesme yoluyla yıkım ile sonuçlanır.[13]
Küçük müdaheleci RNA (siRNA)
değiştirMetazoanlarda, Dicer tarafından işlenmiş küçük çift iplikli bir RNA türü, RNA uyarımlı susturma kompleksi (RNA-induced silencing complex, RISC) içine alınır. Bu komplekste yer alan bir endonükleaz, bu küçük RNA ile baz çifti kurabilen mRNA'ları parçalar. siRNA, çift iplikli RNA virüslerine kaşı bir savunma mekanizmasıdır. siRNA ayrıca, laboratuvarda, hücre kültüründe genlerin fonksiyonunu bastırmak için kullanılır.[14]
Mikro RNA (miRNA)
değiştirMikroRNA (MiRNA), mesajcı RNA'nın dizisine neredeyse tam komplemanter olan kısa RNA'lardır. miRNA'nın mRNA'ya bağlanması sonucu mesajın çevrimi baskılanabilir veya başlık çıkarma kompleksi tarafında 5' başlık çıkartılabilir.[15] miRNA'nın çalışma mekanizması aktif olarak araştırılmaktadır.[16]
Diğer yıkım mekanizmaları
değiştirMesajların yıkımına neden olan başka mekanizmalar da vardır, bunların arasında bitişsiz yıkım (İng. Non-stop decay), Piwi-etkileşimli RNA tarafından susturma sayılabilir.
Kaynakça
değiştir- ^ Job, C.; Eberwine, J. (1912), "Localization and translation of mRNA in dendrites and axons" (w), Nat Rev Neurosci, cilt 2001, ss. 889-98, 1 Haziran 2012 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Ainger, Kevin; Avossa, Daniela; Diana, Amy S.; Barry, Christopher; Barbarese, Elisa; Carson, John H. (1997), "Transport and Localization Elements in Myelin Basic Protein mRNA", The Journal of Cell Biology, 138 (5), ss. 1077-1087, doi:10.1083/jcb.138.5.1077, PMID 9281585, 28 Kasım 2007 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Shabalina SA, Ogurtsov AY, Spiridonov NA (2006). "A periodic pattern of mRNA secondary structure created by the genetic code". Nucleic Acids Res. 34 (8). ss. 2428-37. doi:10.1093/nar/gkl287. PMC 1458515 $2. PMID 16682450.
- ^ Katz L, Burge CB (Eylül 2003). "Widespread selection for local RNA secondary structure in coding regions of bacterial genes". Genome Res. 13 (9). ss. 2042-51. doi:10.1101/gr.1257503. PMC 403678 $2. PMID 12952875.
- ^ a b Kozak, M. (Mart 1983). "Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles" (PDF). Microbiological Reviews. 47 (1). ss. 1-45. PMID 6343825. Erişim tarihi: 12 Ağustos 2006.[ölü/kırık bağlantı]
- ^ Wells, S.E.; Hillner, P.E.; Vale, R.D.; Sachs, A.B. (1998), "Circularization of mRNA by Eukaryotic Translation Initiation Factors" (w), Molecular Cell, 2 (1), ss. 135-140, doi:10.1016/S1097-2765(00)80122-7[ölü/kırık bağlantı]
- ^ Deana, Atilio; Celesnik, Helena; Belasco, Joel G. (2008), "The bacterial enzyme RppH triggers messenger RNA degradation by 5", Nature, 451 (7176), s. 355, doi:10.1038/nature06475, 21 Ocak 2008 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Parker, R.; Sheth, U. (2007), "P Bodies and the Control of mRNA Translation and Degradation" (w), Molecular Cell, 25 (5), ss. 635-646, doi:10.1016/j.molcel.2007.02.011, 20 Ekim 2018 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Shaw G, Kamen R (Ağustos 1986). "A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation". Cell. 46 (5). ss. 659-67. doi:10.1016/0092-8674(86)90341-7. PMID 3488815.
- ^ Chen, C.Y.A.; Shyu, A.B. (1995), "AU-rich elements: characterization and importance in mRNA degradation", Trends in Biochemical Sciences, 20 (11), ss. 465-470, doi:10.1016/S0968-0004(00)89102-1, 23 Kasım 2017 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Chen, C.Y.; Gherzi, R.; Ong, S.E.; Chan, E.L.; Raijmakers, R.; Pruijn, G.J.M.; Stoecklin, G.; Moroni, C.; Mann, M.; Karin, M. (2001), "AU Binding Proteins Recruit the Exosome to Degrade ARE-Containing mRNAs", Cell, 107 (4), ss. 451-464, doi:10.1016/S0092-8674(01)00578-5, 17 Eylül 2011 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Fenger-grvrn, M.; Fillman, C.; Norrild, B.; Lykke-andersen, J. (2005), "Multiple Processing Body Factors and the ARE Binding Protein TTP Activate mRNA Decapping" (PDF), Molecular Cell, 20 (6), ss. 905-915, doi:10.1016/j.molcel.2005.10.031, 6 Haziran 2011 tarihinde kaynağından (PDF) arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Isken, O.; Maquat, L.E. (2007), "Quality control of eukaryotic mRNA: safeguarding cells from abnormal mRNA function", Genes & Development, 21 (15), s. 1833, 16 Mayıs 2008 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Obbard, D.J.; Gordon, K.H.J.; Buck, A.H.; Jiggins, F.M. (2009), "Review. The evolution of RNAi as a defence against viruses and transposable elements", Philosophical Transactions of the Royal Society B: Biological Sciences, 364 (1513), s. 99, doi:10.1098/rstb.2008.0168, 1 Ağustos 2013 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Rehwinkel, J.A.N.; Behm-ansmant, I.; Gatfield, D.; Izaurralde, E. (2005), "A crucial role for GW182 and the DCP1: DCP2 decapping complex in miRNA-mediated gene silencing", RNA, 11 (11), ss. 1640-1647, doi:10.1261/rna.2191905, 5 Haziran 2008 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
- ^ Eulalio, A.; Huntzinger, E.; Nishihara, T.; Rehwinkel, J.; Fauser, M.; Izaurralde, E. (2009), "Deadenylation is a widespread effect of miRNA regulation", RNA, 15 (1), s. 21, doi:10.1261/rna.1399509, 1 Ağustos 2013 tarihinde kaynağından arşivlendi, erişim tarihi: 6 Şubat 2009
Dış bağlantılar
değiştir- mRNA ömrü 27 Eylül 2007 tarihinde Wayback Machine sitesinde arşivlendi. animasyon